|
EVIDENCE OF THE
PRECISION OF
GENETIC ADAPTATION
by
H. J. Muller
Lecture delivered February 19th 1948.
Published in Harvey
Lectures, 43, 165-229
(With
copyright permission from Academic Press. Note that "footnotes" are by HJM, and
"commentaries" are by DRF.)
OPPOSITION TO ADAPTATIONIST
INTERPRETATIONS
MUTATION
AS NATURE'S DEMONSTRATIONS OF THE USEFULNESS OF NORMAL CHARACTERS
NORMAL
GENES ARE NEAR THE SATURATION LEVEL OF THEIR EFFECTIVENESS
DIFFERENCES
IN GENE DOSAGE THAT EXIST BETWEEN THE SEXES
DOSAGE
COMPENSATION AS A NORMAL ADAPTATION
THE SURPASSING
PRECISION OF ADJUSTMENT THEREBY IMPLIED
AN
OBJECTION BASED ON CORRELATION BETWEEN USELESS AND USEFUL CHARACTERS
THE
DEVELOPMENTAL MECHANISMS INVOLVED IN DOSAGE COMPENSATION
"SECOND-ORDER"
PRESSURE OF SELECTION IN ESTABLISHING DOSAGE COMPENSATION
ADAPTIVE
PRECISION BASED ON THE PROLONGED PERSISTENCE OF MINUTE FEATURES
INDIVIDUAL
VARIABILITY AS A MEASURE OF THE LATITUDE OF ADAPTIVENESS
SUCCESSES AND LIMITATIONS IN
ADAPTATIONS
SUMMARY
THE OPPOSITION TO ADAPTATIONIST
INTERPRETATIONS
ALTHOUGH the present theme is a general biological one, drawing mainly
upon data supplied by so insignificant an object as the fruit fly, Drosophila
melanogaster, nevertheless the problems involved are so fundamental that they bear as
strongly upon medical matters as upon those concerned with other organisms than man.
To be sure, no
medical man, familiar as he is with the stupendous intricacies of bodily structure
and function, would deny the marvelous complexities of adaptation attained by the human
organism, both in their chemical, physiological and morphological aspects. Yet it has for
almost half a century been a favorite attitude, among a number of schools of
physiologists, and of those general biologists in fields related to physiology, to regard
the search for functions of discovered structures, processes or substances of organisms as
an old-fashioned and philosophically erroneous pursuit, a relic of the vitalistic outlook.
We should, they feel, be satisfied with looking for the cause of the given phenomenon in
data of biochemistry, of ontogeny, and perhaps too of variation, and while we may also
look for the effects produced by it, we should shun the thought of values, or the search
for the functional in it, for that would be teleological and essentially mystical. The
ground for teleology, they would say, has been cut away from under modern biology,
physiology and medicine by the success of the physicochemical attack. Similarly, even
among many of those biologists who are further removed from medical fields, there has long
been a grave distrust of interpretations based upon the usefulness of characters.
Whatever its outer
dressing, this tendency stems for the most part from that reaction
against Darwin's theory of the survival of the fittest as the guiding factor in evolution
which had its heyday during the several decades just before and just after the turn of the
century. Among paleontologists, for instance, there are those who still believe in evolution through a sort of inner compulsion or orthogenesis,
holding that some lines become "senile," or undergo a forced overdevelopment in
certain directions, to their own disadvantage, before dying out.
But what has
seemed the most favorable field for anti-selectionist,
anti-adaptationist views has been that of apparently unimportant structures, such
as a demonstrably dispensable appendix, and of small differences between related groups,
such as a larger horn on one species of beetle, or a more divided leaf on some sub-species
of tree. Not a few biologists, wishing to get away from such minutiae of selection, and to
throw the blame for things they cannot understand on chance, would deny
selective values here and restrict them chiefly to the more intelligible
differences between the broader categories of organisms, such as orders, classes and
phyla.
An especially fine
road of escape from all petty selection seemed a half-century and less ago to be provided
by de Vries's theory of the origination of so-called "elementary
species" by single "great mutations,"
involving many characters radically at one bound. Now that the very special nature of most
of the observed de Vriesian "mutations" is
understood, a bid has been made for the acknowledgment of some other grand process of
genetic reorganization, involving radical chromosome changes
or "mass mutation," as the major means of
origination of species. However, the data have failed to convince many geneticists of this
conclusion.
Much better founded,
on the other hand, is another neo-Mendelian trend. This starts out with the recognition:
First, of the random, non-adaptive character of individual mutations in
general.
Second, of the fact, so justly
stressed by Wright, that some of these mutations will become multiplied and established in
a population through a purely accidental process-"drift."
As a result of this combination of
chance circumstances it turns out, according to some critics of natural selection, that
many, and perhaps most, of the features seen to distinguish species, and even some broader
categories, are non-adaptive. And, whether or not this fortuitous establishment of
characteristics is accepted, it is still more widely asserted that there is another
considerable group of useless traits which have become incorporated in the bodily make-up
simply because they happened to be by-products of some useful processes or structures,
necessary concomitants in their development, which became automatically included in the
picture as a result of the really important character having been actually selected for.
This latter view is in fact to be found even in Darwin's Origin
of Species itself.
No such broad
reappraisal of the processes of evolution, as the above claims would seem to call for, can
be attempted in our restricted space. We will limit ourselves here to the consideration of
a few related lines of attack on these questions, afforded by mutation study. These have
not often enough been thought of in connection with the present subject, but it is to be
hoped that they may hereafter be given more attention. For these lines of work, although
begun some time ago, have hardly been prosecuted as yet to the point of diminishing
returns, and they are probably capable of a good deal more development. And although the examples which will be cited in this field must for the
present be confined mainly to Drosophila, since that is the material in which most of the
work along these lines has been carried out, and with which also the writer happens to be
the most familiar, it seems very unlikely, in view of the nature of the evidence, that
their significance is limited to this organism. |
|
[Commentary:
Muller here notes "anti-adaptationist", "anti-selectionist" schools, which appear to oppose classical Darwinian
theory based on Natural Selection for "survival of the fittest". However, among these, the school stressing the role of random Variation
and Drift is held to be "much
better founded". Studies of
mutations in Drosophila bear on the controversy, having a significance unlikely to be
"limited to this organism", but "have not often enough been thought of" in this context.]
|
MUTATION
AS NATURE'S DEMONSTRATIONS OF THE USEFULNESS OF NORMAL CHARACTERS
It is surprising how little it has been generally realized what
excellent objects mutations afford for the controlled study -- by contrast with them -- of
the functions of members and features normal to the organism.
The gene change provides us with a subtle inner operation which is often capable of
excising, remolding or changing the properties of an organ, tissue, cellular component or
substance, far more deftly than this could he done by the surgeon's or experimental
physiologist's knife or other relatively crude procedure, or even by the pharmacologist's
or biochemist's medication.
It has, to be sure,
been evident for some years to some geneticists with physiological and embryological
leanings what rich opportunities visible and other mutations offer for the investigation
of the ontogenetic, the physiological, and ultimately the biochemical processes whereby
the observed characters are brought into being, and a few exemplary researches have been
made along such lines, in a number of different organisms.
But it has not
been so well appreciated how valuable this material also is for investigations of the
functions of the given characters, in the individual's living and perpetuation (although
of course these two lines of study are closely connected and in fact not ultimately
separable). For this reason my examples of conclusions that may be drawn in this way in
Drosophila are derived mainly from the results of casual inspection of easily visible
mutants, and seldom upon researches carefully directed to these ends. Nevertheless, even
the incidental observations have already been sufficient to show the distinct trend of the
data, with reference to the chief question here at issue.
Let us first recall,
by a scrutiny of our illustrations of normal male and female Drosophila (Fig. 1), how very
complicated even this external anatomy is, how many details it has which possess no value
obvious to us, but how nearly constant, on the whole, most of these features are from
individual to individual, so that every bristle, every row of hairs, and every veinlet of
the wings, has been given its name or number.
Yet, recall also that
by careful search, mutations changing or abolishing one or another of each of these
characters to a greater or lesser degree, and either singly or in some combination, may be
and have been found, and that there are probably still more frequent
mutations which alter them so slightly as not to have been noticed. Undoubtedly the
same story could be repeated in every higher organism. Hitherto, however, only the
mutations with more conspicuous effects have been much worked with, on account of the
greater ease with which they are recognized, and so our examples will be confined to these
for the present. |
|
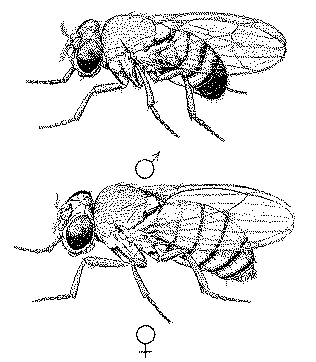
|
| FIG.
1. Drosophila melanogaster, normal type, male above, female below |
If we
now turn our attention to the fly's most conspicuous appendage, the wings, examining the
wing mutations from their functional aspect, we find, as we should expect, that those
mutants which do not fold their wings back completely while these are at rest are apt to
get them caught and bedraggled. That is, the normal position of the wings is more
advantageous. So is the normal shape and size, as shown by the fact that those with wings
abnormal in shape or unusually small are visibly hampered in flying. We may by the way
surmise that, conversely, oversize wings also are less efficient, in relation to the
weight of the fly and to the energetics of the muscular mechanism for beating them. The
many-millions-of-years-old pattern of the wing veins has been by no means fortuitously
fixed upon either. Thus, if even a little cross-vein is absent, or a portion of a long
vein, the wing is unduly flimsy, and likely to become torn. Moreover, branched veins
having blind ends to their branches are likely to leak fluid into the wing at these
points, resulting in blisters or even ballooning.
The bristles and
hairs afford another set of external features in which conspicuous mutations occur. They
cannot be there for warmth in this tiny object, or for their pleasing appearance. The
presence of any particular one in its regularly assigned place might seem to be a matter
of no importance. Yet an inspection of cultures of given mutants, in which particular
bristles are absent or reduced in size, shows that such a seemingly trivial deficiency is
apt to cause trouble. For such a tiny, weak creature as an insect the capillarity of water
and the adhesiveness of the moist soft materials with which it must often come into
contact constitute an extremely powerful force and a serious menace. The waxed hairs and
bristles tend to keep this danger at a distance and flies lacking them are much oftener
found stuck in the food or on any moist surface. Even where one small group of bristles is
absent, as in Fig. 2b showing the so-called "scute"
flies, which chiefly lack the bristles on the small scale or "scutellum" that
forms the posterior end of the thorax, we may frequently see that a bit of the viscous
culture medium has become plastered upon them in just that region. |
The same disasters are likely to meet flies which have bristles or hairs that are
unusually small, or that are split and curled, like the so-called "forked" bristle flies of Fig. 3, or that are otherwise
misshapen. Similarly, it has been shown, mutants with those chemical changes in their
cuticle that go with the color change to "yellow",
and to certain other types of pigmentation, are less resistant to desiccation (Kalmus), as
well as more subject to getting glued up. |

|
| FIG.
2. Photograph of two Drosophila melanogaster females. Fly above is of the
"scute" type, lacking bristles on the scutellum, while fly below is normal in
this respect (note its large bristles projecting dorsally from the middle of the back).
Both flies have narrow eyes of the mutant type called "Bar" (in this case the
less extreme Bar type in which the abnormality is heterozygous, i.e., derived from one
parent only). |
 |
An
additional danger, for all flies defectively equipped with bristles or hairs, is that on
them their external parasites, the mites, have a much easier job in becoming attached, as
they do, to the epidermis, so that one commonly sees these flies much more heavily
infested. |
| FIG. 3.
Drosophila (male) with "forked" bristles (the chief visible effect being their
crinkliness). |
A last example of
bristle function may be drawn from the minute pair of hooked bristles at the end of each
foot. For the mutants in which these are underdeveloped (such as those which, by a second
effect of the same gene, have "spectacled" eyes)
are, unlike the normals, incapable of getting a good grip on vertical or inverted surfaces
and walking readily upon them.
It is widely known
that the normal brick-red eye color in
Drosophila is produced by the cooperation of dozens of genes and that most of their
mutations result in the pigmentation being lighter. Can it be by mere chance that this
complicated outfit for keeping the color dark continues to be maintained? Although even
the white eye is sensitive to light, and can distinguish the general direction of its
source, tests by Kalmus and others have shown that light colored eyes do not enable the
flies to orient as well to moving patterns (stripes). This is presumably because in them
the light is more indiscriminately diffused over the photosensitive retinal cells, rather
than in the form of the sharp, erect images produced by the light that passes through the
well-pigmented light-insulated optic tubes of the normal compound eye.
Now, the same genes
which give rise to this optic pigmentation produce pigmentation also in some internal
organs, notably the excretory tubules and the sheath of the testis. And tests of Mackenzie
and this writer have shown that flies without this screen over their gonads, thin though
it may be, are considerably more sensitive than the normal to the damaging effect of
ultraviolet light (of wave lengths present to some extent in outdoor sunlight) in
producing mutations in their germ cells. The normal epidermal pigment likewise helps in
this respect.
The above
observations, superficial though most of them have been, will suffice for our present
purposes. Many other externally visible features of little understood function, such as
the three small ocelli, the parts of the antennae, the individual joints of the legs,
projecting mouth parts, secondary sexual characters, and characteristics of the behavior,
could be studied in a similar way, and would in fact justify much more intensive studies.
At the same time, the interior characteristics of the body, including especially those
pertaining to the cells themselves, and to their metabolism, remain as yet a largely
unread library.
While few other types
of visible mutations than those above discussed have been studied enough to show their modus
operandi in affecting the life of the organism, one general fact concerning them
deserves emphasis here. This is that practically all of them do result in some
quantitatively demonstrable reduction in the expectation of life, even under the
relatively sheltered conditions of laboratory culturing. In many of these cases, to be
sure, it is evident that there must be some hidden damage, besides that stemming from the
visible change itself, inasmuch as even the larva, in which the given external
feature has not yet had a chance to develop, is less viable.
This illustrates the
multiple effects which most individual genes have. We have however intentionally chosen,
for our above examples, cases in which the consequences of those changes that occurred in
the visible characters themselves could be traced. For only thereby was it possible to
show that all these characters too, non-essential though they might seem, did have their
own utilitarian roles to play in the great system of processes that serves biological
perpetuation.
At the same time it
should be remembered that all so-called visible mutations together, including those which
have hidden harmful effects accompanying them, form only a small minority -- at least as
low as a twentieth -- of all the mutations that can be demonstrated to occur in
Drosophila, and that the vast majority of all these changes is somehow detrimental, just
as one would expect for fortuitous alterations in processes that themselves play
some serviceable role in the organic system.
That this is true on
the biochemical level is clearly evident from the work of Beadle and his co-workers on
moulds, and this work also demonstrates the value of such mutations for the elucidation of
the biochemical functions of the genes there studied, just as in the Drosophila cases
above cited the functions of the morphological characters are brought to light.
In passing we may
note, for the benefit of possible objectors here, that there is no real contradiction
between mutations having provided the building blocks of evolution, on the one hand, and
the fact of such a large majority of them being detrimental, on the other hand. For the
very fact of the organism's having been fashioned largely through a step-by-step natural
selection of those rare mutations which did happen to fit, carried on over many millions
of years, would necessarily have resulted in its being so organized now that a random
alteration in almost any feature would be far more likely to be harmful than helpful to
it.
This could still
leave open, however, various devices and special routes for its further improvement,
especially those which fitted it to hitherto unusual conditions, and it is evident that
upsets of the organic balance in any such directions would in turn be likely to open up
still further possibilities. Thus only the very rare mutation, preferably too under rare
conditions or in rare combinations with other mutations, can be expected to be useful in
evolution, if in fact organisms have evolved through an accumulation of useful mutations.
And, as we have seen, the data on the usefulness of existing characteristics quite fit in
with this interpretation.
It might however
still be maintained that, although the majority of organic features do have their useful
functions, evidence of the above kind indicates this only in a rough way, and that we are
still at liberty to hold that alterations in somewhat less conspicuous characters, or
finer grades of change than those studied in the above characters, would be of
indifference so far as the organism's success in living was concerned. It is thus evident
that more exact methods, or a quite new mode of attack, would be desirable for throwing
light on the question of the adaptive value of character differences of a more minute
order of magnitude. Fortunately, evidence of an unexpected nature has in fact turned up,
in Drosophila genetics of a much more technical kind, concerning the
adaptiveness of differences of an astonishing degree of refinement, even in cases
where the reason for the functional importance of these differences is not known. |
|
[Commentary: Muller here notes
that: "the vast majority" of mutational changes are "detrimental, just as one would expect for
fortuitous alterations in processes that themselves play some serviceable role in the
organic system", and
that: "practically all
of them do result in some quantitatively demonstrable reduction in the expectation
of life". However:
"only the mutations with
more conspicuous effects have been much worked with, on account of the greater ease with
which they are recognized".
Some
characteristics of various mutants are noted. To what extent do "alterations in somewhat less conspicuous
characters, or finer grades of change" affect "the
organism's success in living?"
"Evidence of an
unexpected nature" now
throws "light on the
question of the adaptive value of character differences of a more minute order of
magnitude".] |
EVIDENCE
BASED ON THE FINDING THAT NORMAL GENES ARE NEAR THE SATURATION LEVEL OF THEIR
EFFECTIVENESS
This evidence arose out of studies of the observable effect on the
organism produced by changing the quantity of given genes.
For this purpose, comparisons were made of individuals having the normal number, or "dose", of these genes, with those which were
known to have inherited more or fewer than the normal number, or dose. Such differences in
dose were made possible by first irradiating the germ cells of individuals having the
desired genes and then, in appropriate crosses, picking out and breeding -- for extra
doses -- those offspring which happened to have received a small extra fragment of
chromosome, broken in just the right way to contain that gene, or -- for subnormal doses
– those offspring which received all of the chromosome in question except for a
corresponding fragment which in this case had been broken out of it and lost.
An illustration of
the most typical sort of situation is given in Fig. 4, which shows the results with the
sex-linked mutant gene for apricot eye color. This is a rather light color produced by a
mutation of the same normal gene which, by a more extreme mutation, sometimes gives rise
instead to the well known white eye. Let us confine our attention at first, for
simplicity's sake, to the results in females. |
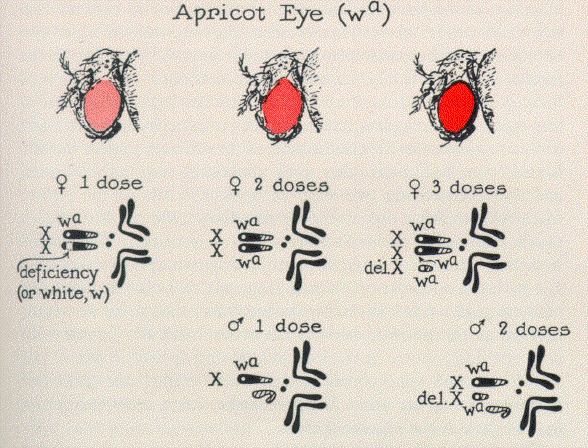
FIG. 4.
The apricot eye mutant of Drosophila as expressed in flies of different genetic types. The
gene for apricot is represented by the symbol wa. The term "del. X"
denotes a deleted X chromosome, that is, one which has all the middle
portion gone and consists only of the extreme left and right end portions joined together;
the gene wa is present in the left hand portion of this deleted X.
The
chromosome constitution of a series of these is represented on the upper row of
cytological diagrams in this figure. The diagram in the second position from the left end
of this row indicates the constitution of the ordinary apricot female, having two X-chromosomes
each containing one apricot gene, and therefore having two doses of apricot in all.
As the picture
of an eye just above this on the top row shows, the resulting hue is rather like that of
the fruit after which it is named. The female with three doses, to the right of it, having
two apricot-containing X's with the addition of a small fragment that
also includes one of these genes, has a distinctly darker color, though by no means as
dark as the normal red. On the other hand the female with only one dose, shown at the left
end, having apricot in one of her X-chromosomes but a small section which
could have contained it missing from her other X-chromosome, has a color
only about half as dark as the regular apricot.
The above comparison
shows, for one thing, that the amount of pigment varies with the dosage, and in a nearly
proportionate manner in these cases. In this connection it may be mentioned that, as a
more refined check on the conclusion that it was really the change in dosage of the gene
for apricot itself which was responsible for the color differences, and not (which seemed
but a remote possibility) that of some other genes which happened to be in the same small
chromosome fragment, the effect of adding a slightly smaller fragment, taken from the same
region but not containing the locus of apricot, was tested out. This effect was found, as
expected, to be zero.
A second conclusion
to be drawn from the results is that, since the observed color
varies directly rather than inversely with the dosage, the gene which we call
"the gene for apricot" must be in some way engaged
in furthering rather than in interfering with the production of eye pigment. However, it
is not as effective in its work of aiding in pigment production as is the normal gene,
since the presence of two or even of three doses of the gene for apricot fails to lead to
the intense brick-red coloration found in normal individuals.
Before such
dosage tests had been made this conclusion could not yet legitimately be drawn. For it
might as well have been assumed, as an alternative, that the mutant gene called apricot
exerts some actual inhibiting action on the process of pigment manufacture, which normal
genes simultaneously present in other positions in the chromosomes are working to carry
out, and it might even have been postulated, further, that the normal gene from which
apricot arose by mutation (its normal "allele", as
we say) itself played little or no positive role in the pigmentation process (or even a
somewhat negative one). Since however both the normal and the
apricot genes alike are shown by the above seriation to play a positive role, with that of
the apricot similar to but less potent than that of the normal, we have applied the term
hypomorphic here
to designate the relationship of the mutant to the normal gene's type of activity.
It must be understood
that this terminology does not imply that the mutant gene's action is qualitatively, or in
a biochemical sense, exactly the same as that of the normal allele, only less. In fact,
there are several good reasons for inferring that the mutation caused a change in the
chemical constitution of the gene and of its product. What is implied is only that the
mutant gene works in such a way as to produce a final
effect similar to that of the normal, but a lesser effect, like that
which would presumably have been brought about if the concentration or the activity of the
normal gene itself (or of its products) had been reduced. It is in this sense that the
mutant gene is less active.
Results similar
to those above described for apricot have been obtained with a good many other mutant
genes in Drosophila, both with those in the X and in other chromosomes.
We now have available for conclusions in this field not only the studies on extra doses,
which were the first done expressly for this purpose, but also the much more numerous,
still earlier found cases of so-called "deficiencies",
that had been investigated especially by Bridges and by Mohr. These could not with
confidence be used for such conclusions until -- long after their finding -- it was shown
by cytological study of salivary gland chromosomes that a small bit of chromosome is in
fact really absent, rather than altered in some way, in such cases, and that their use
accordingly gives us a means of changing the number of genes present.
Now, a perusal of all
the available cases involving either extra or missing doses of mutant genes, or both,
shows clearly that the great majority of them are, like apricot, hypomorphs.
That is, they have an action like, but lesser than, that of their
normal allele, and varying in amount directly with their dosage. This is true
for instance of the scute gene, previously referred to, as shown by observations on
bristle number in the presence of one, two and three doses, and it is true of the forked
gene, as shown by the frequency and amount of bristle deformation.
There are some mutant
genes, however, like that for white eye, the action of which on the observed character
turns out to be so very weak, if present at all, as to be indetectible, as judged by the
ineffectiveness of change in their dosage. These latter are accordingly termed "amorphs", and may be regarded as standing at
the bottom of the ladder of hypomorphism. On the other hand, there are probably a few, to
be termed "hypermorphs", with even
greater effectiveness than normal, though these are hard to detect for technical reasons.
And there are certainly some, though rare, which we call "neomorphs",
with a qualitatively different type of effect than the normal.* [Footnote: Possibly too there is a rare class of
mutants, opposite in type of action to their normal alleles, that may be termed
"antimorphs". However, our earlier criteria for these were inadequate. For, as
C. Stern has shown, the property of competition between mutant and normal genes is
separate from that of the direction of their effects. Thus the latter must be judged only
from experiments in which the dosage of one allele is varied in the total absence of the
other, a procedure which has been insufficiently tried in the cases in question. This
competition, it may be pointed out, may be either for substrates, as Stern has postulated,
or, conceivably, may involve an interference of some of the gene's products with the total
gene activity of the two alleles.] Although these
last are of particular importance in their bearing on evolutionary possibilities, the
point of greater immediate interest for us in connection with our present theme is that
the great majority of mutations turns out on these tests to be hypomorphic or amorphic;
that is, they involve some sort of weakenings or losses of biochemical processes normal to
the organism, processes the strength of which varies directly with gene dosage.
These results acquire
a greater interest when considered in connection with those for the normal alleles of
these same hypomorphic mutants. In the great majority of cases a change either from two
down to one or up to three doses of the normal gene results in no detectable change in the
character, or at most in a change of trifling magnitude, seldom noticed except under
special conditions. Yet we have just seen that the effect of these normal genes must be
regarded as like that of a quantitatively greater amount of mutant genes.
It must therefore be
concluded that, with increase in the dose (or with an equivalent increase in the activity)
of the genes at these loci, although there is at first, at the lower levels represented by
the hypomorphic mutants, a proportionate rise in the effect, as shown in the left hand
portion of the curve in Fig. 5, there ensues a progressively lesser rise, that is, the
curve levels off in its right hand portion (that representing the grades of effect given
by the normal alleles), until it becomes nearly horizontal. In this right hand portion,
then, the so-called "saturation effect" is
approached. It is not surprising that gene effects, like so many others, should at
high doses obey this "law of diminishing returns." |

FIG. 5. The
relation between eye color and gene dosage or activity in the case of apricot and its
alleles, white and normal. The symbol w+ represents the normal allele of
apricot; wa, as before, represents the apricot allele.
We may
however ask, why is it that the normal genes should in such a
majority of cases be at a level of effectiveness which puts them in this "saturation" portion of the curve?
The answer to this would at the same time go far towards explaining the observed fact that
such a majority of normals appear almost completely dominant to their mutant alleles. For
the finding that most mutants are hypomorphs or amorphs would lead to the conclusion that
the heterozygous individual, i.e. that having one mutant and one normal allele, has the
equivalent of one or somewhat more than one dose of the normal gene, and this, being at or
near the saturation level, would have an effect practically equal to two normals; thus the
normal would appear dominant.* [Footnote: We are here assuming that the mutant gene's competitive action,
mentioned in the preceding footnote, is usually a minor one, as the facts indicate it to
be. In the relatively few cases in which it is more pronounced, however, it must cause the
level of activity in the heterozygote to be lower, approaching more nearly, or even
going appreciably below, the level of one dose of the normal gene.]
In seeking an
interpretation of this high effectiveness of the normal gene we may discount the
possibility that the most advantageous level of a character can only be attained by, so to
speak, pushing each gene to its utmost activity. There is no a priori reason why
characters in general should represent the maximum biochemical possibilities. The body
pigmentation, for example, is not the darkest chemically possible, nor is even that
of the eye, since the normal types of some Drosophila species, as well as some mutant
types of Drosophila melanogaster itself, have darker bodies, and some have
darker (though perhaps not redder) eyes. That is, higher levels are possible by
readjusting other interacting gene processes, and the present lower level might then be
reattained by lowering the effectiveness of the particular gene we are primarily
considering. Thus we should be able to obtain a substitute normal type, which would in the
main be indistinguishable from our present one.
We are also very
skeptical of the thesis that the high potency of the normal has been attained, as Fisher
has suggested, by reason of the advantageousness of having heterozygotes (individuals with
one normal and one mutant gene) rendered nearly normal. For, as Wright and the author have
separately pointed out, such heterozygotes are so few as hardly to give a selective
advantage great enough to withstand the disruptive tendencies of mutation pressure, drift,
and selection for more important correlated characters.* [Footnote: Even if Fisher's explanation were adopted,
however, it would lead us to the same conclusion concerning the high adaptive value of
precisely the "normal' degree of gene expression now existing, as that arrived at on
our own interpretation.]
The clue to an
interpretation not subject to these difficulties is (as was pointed out independently by
Plunkett and the present writer in 1932) provided by the fact that mutant genes are
on the whole much more variable in their expression than normal genes. This is obviously
related to the further fact that most of these mutant genes
(the hypomorphs and the neomorphs as well) have a level of
effectiveness, in regard to the characters studied, such as to place them on the obliquely
sloping, left-hand portion of the previously discussed curve.
For, since they
occupy this position, any agency which operates, during the
time of their activity in determining the character, in such a way as, in effect, somewhat
to diminish or increase this activity, will result in a visible alteration of the
character, for it will have an effect 1ike that of changing the dose.
Agencies, both of an environmental and of a genetic kind, which exert such influences, are
quite evidently very common, as we can see from the character's variability.
Yet, if we were to
judge from the relative invariability of the normal type, these influences are not
at all common. The reason for this apparent discrepancy clearly is that the normal gene has such a high level of effectiveness that even when its
activity (along the abscissa of our curve) is shifted backwards or forwards by the given
influences as much, relatively * (Footnote: i.e. when the variations in activity are expressed as fractions
of the mean activity.), as
that of the mutant gene was found to be (a process which we have no reason to
doubt), there results no discernible alteration of the character
(the ordinate) since the curve of effect is here almost horizontal (see Fig. 5).
But this
conclusion in itself furnishes us, ready made, with the explanation of why the normal gene
has been given this nearly saturation level of activity. It is because variations in the
character exhibited by the normal individual are thereby minimized. All this implies as
its basic proposition that such variations are disadvantageous. That is, that the precise grade ordinarily attained by the normal character must, under
most circumstances, be optimal, and even small deviations from it must result in an
appreciable lowering of the individual's expectation of life or reproductive rate.
When we here say "appreciable" we mean, great
enough to afford an effective handle for the operation of natural selection.
We can even get some
idea of the amount of character alteration which affects survival enough to be of
importance in evolution, on the basis of these considerations. This may be done as
follows. Let us first measure the amount of phenotypic (i.e. visible) variation of a
hypomorphic character, such as apricot, taking, say, its variance under natural conditions
as our index. We may then determine, from our curve of the variation of the character with
gene activity or dosage (Fig. 5.), how great a change in activity, measured on the
abscissa, this observed change in character, measured on the ordinate, corresponds to. Let
us then divide this "activity variance" by the
value for the average activity in the case of apricot itself, so as to put the variance in
relative terms, constituting a "coefficient of variance of
activity" for the apricot gene.
Next, assume
that a like variation in activity of the normal gene, relative to its own value, occurs as
a result of the same fluctuating influences. We may then see how much absolute variation
in activity this amount of relative variation would correspond to in the case of two doses
of the normal gene. After plotting the resulting values for this in their appropriate
positions on the abscissa, on either side of the value for the average activity of two
doses of the normal gene, we may then read off on the corresponding
ordinates how much alteration in the character ("phenotype")
of individuals provided with two normal genes would thereby result.
It will certainly be found that in the given case this amount of variation is quite
imperceptible to our eye. Yet, on this interpretation, a greater amount of variance
than this has been prevented by reason of the disadvantage thereby entailed. In other
words, this quite imperceptible variance represents, in a sense, the limit of the
naturally allowed changes.
It might, as an
alternative, be held that it is not the ordinarily observed variants which are selected
against, but rather the larger ones which may appear under more
extraordinary conditions (environmental and/or genetic). Just
because of the rarity of such conditions we should have to make the selective detriment of
these supposedly larger variations correspondingly higher, for them to be so effective in
selection. But we should thereby arrive at a value similar to that reckoned above
for the average disadvantage of a given amount of change in the character. In any
case, then, we would conclude that extremely high stability has been selected for,
allowing only a subliminal variance.
Summing up the
foregoing argument once more, we may say that the finding that normal genes are in general
so near the saturation level of their effectiveness appears to require the interpretation
that there is usually an appreciable disadvantage to the organism if one of its features
fails to be developed to exactly that grade characteristic of the type which has been
established as normal. In the cases dealt with, in fact, the required exactitude even
surpasses our powers of direct visual discrimination. |
|
[Commentary:
Normal wild-type Drosophilae (w+) have brick-red eyes. Mutants
in eye colour are easily recognized. A null mutant ("amorph") has white eyes. An intermediate sex-chromosome-linked "hypomorph" (wa)
has apricot eyes of a range of increasing intensities corresponding to the number of gene
doses. Fig. 4 shows for each intensity the three pairs of autosomes (two pairs large and
one pair small) and the sex chromosomes. A male achieves with one gene dose what requires
two gene doses in the female (discussed in next section of the paper). This constitutes
the basic observation of "dosage compensation".
However, changes in the dosage of the wild-type gene result "in no detectable change in the character". Fig. 5 interprets this in terms of genetic dominancee such that one dose of w+ is
sufficient to produce full colouration ("saturation
effect"). Doubling w+
gene dosage causes no further increase in colour. In contrast, hypomorphs "have a level of effectiveness … such as to
place them on the obliquely sloping, left-hand portion" of the dose-response curve. Here gene dosage is
limiting, thus permitting dosage effects to be observed.
Finally, Muller seeks to explain why the level of expression of the diploid (w+
w+) has evolved to be on the plateau
of the curve so that:
-
(i) Small, frequent,
variations in product concentration would have minimal effect.
-
(ii) Large, rare,
variations would be effectively buffered. Thus, "because
of the rarity ... we should have to make the selective detriment of these supposedly
larger variations correspondingly higher, for them to be effective in selection". This remark is highly relevant to speculations regarding the heat-shock
response, a phenomenon described by Ritossa in 1961 (Forsdyke, 1994; see
below).]
|
EVIDENCE
BASED ON THE FINDING OF GENETIC COMPENSATION MECHANISMS FOR THE DIFFERENCES IN GENE DOSAGE
THAT EXIST BETWEEN THE SEXES
The above evidence of adaptive precision is, however, surpassed by
that arising from a further series of findings of an unexpected nature, concerning the
dosage effects of genes in Drosophila. These surprising phenomena came to light when genes
contained in the X chromosome were considered and a comparison was made
between the dosage effects found in females, which, having two X
chromosomes, regularly have two doses of such genes,
and those found in males, which regularly have one
dose. The relationships found have been denoted as "dosage
compensation".
A typical example of
this is shown by the mutant gene for apricot eye, the dosage effects of which in the
female have already been considered. The lower row of chromosome diagrams in Fig. 4 shows
the genetic composition of the males which have the eye shades shown in corresponding
positions along the topmost row. It is at once apparent:
First, that in the males, as in the
females, the color varies about proportionately with the dose, at
this level of activity.
Secondly, however, it is equally clear
that the color of the ordinary male, with one dose, is very nearly
the same as that of the
ordinary female, with two doses. The one-dose female, then, is considerably lighter than the one-dose male, and the two-dose female,
correspondingly, is lighter than the two-dose male.
These relations are also shown in the
activity-effect curve in Fig. 5, where the bottom line shows the male doses that produce
the effects indicated by the ordinate heights shown on the curve above them. Here the
dotted lines show the approximate latitude of the error possible in the determinations of
color grade. It will be seen that this error is relatively so slight as to justify us in
giving the rule as: one dose in the male has very nearly the
effectiveness of two in the female.
This peculiar sex
alteration in the effect of any given dose of the gene for apricot must clearly be
produced by the interaction, in the pigment-producing process, of other genes than apricot
that differentiate the sexes. That genes in the Y-chromosome of the male
have nothing to do with this matter is not only an a priori likelihood based on
previous evidence of the relatively small influence of the Y in general,
but has been directly demonstrated here by observations on apricot males without any Y and
on apricot females possessing a Y in addition to their two X's,
inasmuch as these individuals are found to have the same color as the regular apricots.
The influence must
therefore be exerted by a gene or genes lying in the X chromosome, at
some other locus or loci than that of apricot itself. This conclusion follows directly
from a comparison of the genetic compositions of any male and female that have the same
dosage of the apricot gene. For example, the genetic difference shown in Fig. 4 between
the regular apricot female, with its two doses, and the darker colored two-dose apricot
male, that has one apricot gene present in an X of normal size and
another in a small extra fragment of an X, lies (if we disregard the Y)
purely in the fact that the female contains not merely this fragment of a second X
but, in addition, all of the remainder of a normally constituted X
chromosome. In this remainder of the X, then, must lie a gene or genes
whose presence in double dose, when the rest of the genetic composition is held constant,
causes the apricot color to be lighter than that developed when they are present only in
single dose -- in fact, just enough lighter so that the two-dose apricot female is brought down to approximately the level of the one-dose
apricot male.
This
interacting gene or genes elsewhere in the X thus have the effect of
compensating almost exactly for that difference in dosage of the gene in question
(apricot) which ordinarily exists between the sexes. It is for this reason that such
modifying genes are called "dosage compensators".
Their action is such as to keep the two sexes alike in their grade of expression of the
given sex-linked character, despite the difference in dosage of the primary gene under
investigation.
Of the many different
mutations that have occurred at the locus of apricot, giving different shades of eye
color, a considerable majority, including both those of grades darker and of grades
lighter than apricot, show the same phenomenon of compensation as apricot itself. A few
however (notably eosin and ivory) show no compensation, the color being virtually
identical in males and females having the same dosage of the gene, even though in these
cases as in the others the color increases with rise in dose within the same sex. We may
suppose either that, through some qualitative chemical change, the process of pigment
formation mediated by them has become insensitive to the action of the normal
compensators, or that they represent more nearly the normal process and that it is the
others which, through a chemical alteration, have come to have effects sensitive to the
compensators. For reasons to be given below, however, the former interpretation, according
to which compensation is a normal phenomenon, appears the only reasonable one.*[Footnote: Direct proof of this has been obtained
since the above was written (see footnote on p. 192)]
That the phenomenon
is, in Drosophila, a general one, and not to be explained by the accidents of a particular
type of mutation, is shown by a scrutiny of the data derived from other sex-linked
mutations. So for example the hypomorph scute, which has been investigated by bristle
counts of males and females with different doses of this gene, brought about by the
addition and subtraction of small fragments produced by irradiation for this purpose, just
as in the case of apricot, gives exactly the same kind of results as apricot. That is, the
two-dose female is much the same in degree of bristle development as the one-dose male and
it has far fewer bristles than the two-dose male.
Various other
hypomorphic sex-linked mutants have been studied similarly, by comparing the effects of
one dose in the male with those of two doses and of one dose in the female (the single
dose being brought about by having a small piece removed by irradiation, from the
appropriate region of one of the X-chromosomes). Included in this study are the loci of
yellow, achaete, facet, singed, garnet, and forked. Of these cases only one, facet, fails
to show an accurate compensation for the difference in dosage between the sexes. In facet,
the one-dose male is less normal than the two-dose female, as expected for incomplete
compensation, although it is not as abnormal as the one-dose female, so that some
compensation does exist. In all the rest, however, the male with its one dose looks very
much like the female with two doses, and much more nearly normal than the female with only
one dose of the gene.
As a final example we
may take the case of Bar eye, shown in Fig. 6, for which quantitative data based on counts
of the number of optic elements, or ommatidia, present were gathered by Offermann working
in collaboration with the present author. This particular allele of Bar, known as
Stone's Bar Bs, had been obtained by Stone as a result of an
irradiation whereby the X-chromosome, already containing Bar, was broken into two pieces,
in such a way that the gene for Bar became included in the right hand fragment. This was
the smaller fragment, and would in fact appear very much smaller, relatively to the other
piece, than in the diagram, if only the genetically active portions of the chromosomes
were shown. |

FIG. 6. The Bar
eye mutant of Drosophila (Stone's Bar allele) as expressed in females and males having
different doses of the Bar and normal alleles.
By
constructing, through appropriate crosses, flies with one or more of these small
fragments, then, the effects, on the size of the eye, of varying the doses of Bs in
both the male and female could be observed. Since in this case the mutant is not a
hypomorph but a neomorph we find that,
unlike what occurs with apricot, scute, etc., the more doses of Bar there are the more
abnormal is the effect, as manifested in reduction of eye width, while on the other hand
changes in the number of doses of the normal locus have no effect on the eye width.
Nevertheless, the size of the eye of the female with two doses of Bar is not very
different from that of the male with one dose of Bar, while much smaller than that of the
female with one dose, and much larger than that of the female with three doses or of the
male with two doses. In other words, there is a strong tendency to dosage compensation in
this case, even though the developmental reaction here, a neomorphic one, must be taking a
somewhat different course, qualitatively, from that in the normal organism, and one not
summative with the latter. The only other sex-linked mutation yet known to be neomorphic,
however, that known as Hairy-wing, fails to show dosage compensation.
Despite the rare
exceptions to, and the comparatively minor imperfections in dosage compensation, the
phenomenon is so general, especially in the case of hypomorpliic mutants, and the
resemblance of one-dose males is so very much closer, on the whole, to two-dose females
than to one-dose females, that it is impossible
to regard it as accidental. We are therefore confronted with the questions:
|
|
[Commentary:
Although there are some exceptions, in general, hypomorphic genes (and one neomorphic gene
mentioned; Fig. 6) exhibit dosage compensation when located on the X-chromosome.
A role of the Y-chromosome in this phenomenon being excluded, it follows
that "the influence must be
exerted by a gene or genes lying on the X-chromosome, at some other locus or loci, other
than that of" the compensated
gene itself.
With a touch of male chauvinism, Muller sees the two dose female as being "brought down" to the level of the one dose male, rather than the
one dose male being brought up to the level of the two dose
female. Thus, he implies his X-linked "modifying genes", or "dosage
compensators", are inhibitory,
so that their "presence in double
dose, when the rest of the genetic composition is held constant, causes the apricot color
to be lighter than that developed when they are present only in single dose". (This has implications for mechanism which will not
be considered here.)
Muller is able to
winnow the relevant from the irrelevant, concluding "that it is impossible to regard it as accidental. We are
therefore confronted with the questions: (1) What has caused
the organism to develop this genetic mechanism? (2) What is its nature
in more detail?" The latter
question (i.e. mechanism) has been the major focus of the succeeding reductionist
generation of dosage compensation researchers.] |
DOSAGE
COMPENSATION AS AN ADAPTATION DEVELOPED IN CONNECTION WITH THE NORMAL TYPE
Turning our attention to the question of the cause of the
phenomenon, we must, to begin with, reckon with the at first sight surprising fact, that
the above cited manifestations of dosage compensation are all exhibited by mutant genes (mainly hypomorphs). It is evident that there could be only the most minimal advantage to the species in having
these mutant genes attain one degree of expression rather than another, in view of
the fact that they occur so rarely in the
population anyway, and that they are especially rare in that homozygous condition in
females in which the compensators exert their characteristic effect.
Moreover, it is
found in the case of the locus of apricot, and of some others in which several grades of
hypomorphic alleles are available, that in general the compensation works as well for one
grade of mutant allele as for another, making the female about equal to the male. Now
there can hardly be an advantage in keeping the apricot female's color as low as it is, if
at the same time there is an advantage, in the case of the darker allele coral, in having
the grade of the female (though here too kept down to that of its own corresponding male)
kept as high as it is in this latter case. The effect on the manifestation of the mutant character must therefore be an
automatic result of the operation of processes the reason for existence of which lies in
their effect in some other connection.
What then is this
"other connection"? There can be but one answer
here. |
|
The
compensators must be genes, established in the normal type, which have the effect of
equalizing the expressions of the normal alleles of the mutant genes which we have
been studying, so as to make the female's two doses of the normal gene produce the same
degree of development of the given character as does the male's one dose of the same
normal gene. And the fact that the phenomenon is so general in its
occurrence, indicates that this result must entail some advantage to the individual and to
the species.
|
This advantage cannot lie in the fact of the resemblance itself, between male and
female, that is thereby brought about in the given characters. Rather it must mean that,
as regards most features and characters, whatever is the best grade
of expression for an individual of one sex is also, and for the same reasons, the best
grade for one of the other sex as well. For after all the males and females of the
same species usually have much more in common, in their needs, way of life, physiology,
ecology, etc., than they diverge in; and, while secondary sexual differences certainly
should and do occur, these are, from the standpoint of the total organismic make-up, a
comparatively minor part of the whole.
But the genes
in the X-chromosome are numbered in the hundreds or even thousands, and
they concern all sorts of bodily processes and structures. Therefore, in the case of the
great majority of them, that grade of development of their effects which is best for one
sex is pretty much the same as the best grade for the other sex. Since, however, the
female has two doses of each such gene and the male but one, the attainment of that same
grade by both would require some special genetic mechanism, one only to be achieved by
what we have termed "dosage compensators".
A converse proof of
the existence of dosage compensation as an adaptation to the end of equalizing the
expression of genes of which the two sexes have unequal doses is provided by an
examination of the dosage effects of non-sex-linked genes. Here too we find, mainly from
studies involving chromosomes with small sections missing, that most mutant genes are
hypomorphs, giving a more normal grade of character as a result of two doses than of one.
But here the two doses in the female give an effect like that of two doses in the male, in
accordance with the fact that here both male and female do regularly have the same dosage.
Thus, the sexes are again usually equal, but, compensators being unnecessary to achieve
this result, genes having the property of compensators do not occur.
A still more crucial
test of the same point is furnished by the case of the hypomorphic mutant called "bobbed," which reduces bristle size, a case worked out by
Stern (1929) before dosage compensation was known. This gene lies in the X-chromosome,
but in that small part of it which has a homologue in the Y-chromosome,
so that, unlike the vast majority of sex-linked genes, the male has the locus represented
in both its sex chromosomes, there being ordinarily one so-called normal allele in the X
and, in effect, at least one* in the Y. [Footnote: It is probably present in full or nearly full strength
in just one arm of the Y (the short arm), and in the form of a weaker
allele in the other arm, as Neuhaus first showed. Thus the male would have it represented
thrice, but its total dose is probably not much, if any, more than double in this sex, so
that the male's dose is not very different from the female's.]
Now the dosage
studies showed that in this case, as with non-sex-linked genes, there was no dosage
compensation, inasmuch as a male with one dose of the gene for bobbed, and lacking a Y,
looked much like a female with one dose of it, while a female with two doses had much
better developed bristles than either. A remarkable check on this case, being in a way the
converse of the converse, is found in the genetic situation with respect to bobbed in the
related Drosophila species simulans, as disclosed by data of Sturtevant's (1929).
Here the Y does not contain an allele of bobbed like the normal allele
present in the X, but contains in some strains an amorph (i.e. fails to
influence the bobbed character), and in others an allele which, like an antimorph, acts to
intensify the abnormality when the mutant gene for bobbed is present in the X.
Thus, dosage
compensation of bobbed would certainly be expected in D. simulans, if it really
exists to serve the end of character equalization. And this is in fact found to be the
case. For, unlike what occurs in D. melanogaster, the simulans bobbed male
without a Y, having its one dose of bobbed in the X,
possesses as well-developed bristles as the female with two doses of bobbed. All these cases, then, combine to show that it is not the fact of a gene's
being in the X-chromosome, per se, which somehow leads to its being subject to
dosage compensation, for this phenomenon appears only in the case of those genes whose
normal alleles are regularly in different doses in the two sexes.
Further evidence that
the chromosome configuration in itself has nothing to do with the matter, is seen in the
cases in which a piece of the X-chromosome has become broken off and
attached to another chromosome and/or, conversely, in which a part of another chromosome
has become translocated onto the X.
Whether the pieces are large or small, or derived from one or another chromosome region,
the result is the same: the genes, both those originally of the X and
those of other chromosomes, still have the same dosage effects as they did in their old
positions. Compensation is a chemical mechanism, or
rather, system of mechanisms, stably established in the distant past,
with reference only to those particular genes which regularly existed in different doses
in the two sexes, and so it continues to operate now even when we change the very
conditions that must once have called it forth.
If our thesis is
correct, dosage compensation is a mechanism normally at work to equalize the expressions
of the different doses of normal genes present in the X-chromosomes
of the two sexes, and it must have become established as a result of the advantage
conferred by this regulation of the expression of the normal genes. Why then have we drawn
our above cases from the effects observed in the case of mutant genes? |
|
Quite obviously this is
because of the mutant genes having a level of
activity so low that differences in their dosages or activity give an easily
observable result, whereas for normal genes, lying as they do
near their saturation level, the difference in expression resulting from two as compared
with one or three doses are usually imperceptible, as has previously been
pointed out. Dosage tests, like those above described for mutants, have in fact
been carried out for the normal alleles of all these mutants. But since the individuals,
whether male or female, with one or two or three doses looked sensibly alike in the great
majority of cases, no conclusion as to whether dosage compensation was or was not
occurring could be directly derived from these observations. The
mutants, in other words, were required by us to serve as indicators, or sensitizers, for
processes and relations which we would not otherwise have been aware of, but which we
cannot avoid concluding must really be present in the normal organism as well.
|
* [Footnote: * More delicate than direct observation
of eye color, however, is electrophotometric determination of the relative concentration
of extracted pigment. R. M. Valencia, F. Verderosa and the author have, since the above
was written, applied this method to the red pigment of eyes having the normal allele
of apricot. The results show unequivocally that this normal allele does in fact (like
apricot but unlike eosin) have dosage compensation, just as had been expected on our
theory presented above. (Footnote added in proof, May 14, 1949.)]
But although the
great majority of normal genes fail to show enough visible dosage effect to make such
studies possible on the normal alleles of most known mutant genes, it would be strange if
the X-chromosome, containing as it does from one to several thousand separable genes in
all, did not include some normal genes that were far enough from their saturation level to
be open to such investigation, either singly or collectively.
That this is true is
demonstrated by the fact that the subtraction of a comparatively small section from one of
the X-chromosomes of an otherwise normal female, though it only changes
the dosage of relatively few of her normal genes, frequently leads to a visible
morphological abnormality (aside from effects on sex and sexual characters), of a type
depending on what region is subtracted. The "deficiency," as it is called, is also
apt to reduce the viability in some way, and it is certain to cause death if the piece
removed is a rather large one, or if it includes any one of a number of particular
loci. These results prove several points. It becomes evident:
First, that there are a few normal
genes whose effect is considerably different in one dose than in two. It is only to be
expected, however, that these seldom happen to be the normal alleles of the relatively few
visible mutations which we have to work with.
Second, it is evident that there are
many normal genes whose effect is slightly different in one than in two doses, so that by the reduction to single dose of many at once the combined
effect becomes perceptible where the individual effect would not be.
Third, the doses that these genes
normally have, and their normal relationships to one another, are the ones most
advantageous for the organism.
Fourth, these normal genes are subject
to dosage compensation.
At the risk of
laboring our argument, it is important that we see clearly how the fourth conclusion above
is arrived at. It depends on the fact that, when we reduce the dosage of the normal genes
in a given section of the X-chromosome from 2 to 1, by removing this
section from one of the two X's of a female, we establish the dosage of these genes which the male normally has. The normal male
however does not exhibit the morphological or physiological abnormalities that appear in
the females with these deficiencies. This means that in it, the normal genes in question,
though present in only one dose, attain about as high an effectiveness as is achieved by
the two doses of them in the female. This difference in effect can only be referred to the
double dose which the females have of genes in other parts of the X-chromosome,
that must act as compensators.
Further evidence of
the same sort of thing is furnished by studies of the effect of adding
sections of the X that contain normal genes. This operation too
results in definite syndromes of visible and physiological abnormalities (aside from
effects on sex and sexual characters), especially if the piece added is large, although,
as might be expected from the shape of our curve of effectiveness (Fig. 5), a larger
addition than subtraction can be tolerated. But in such cases it is always evident that,
while addition of a given piece to a male (leading to two doses of it) results in changes
similar in kind to those produced by the same addition to a female (leading to three doses
of it in her), the changes in the male are considerably more pronounced.
It will be seen that
this male, having thereby been given only two doses of the normal genes in question, has,
so far as these genes alone are concerned, a genetic composition which the female
tolerates with entire impunity. The difference in effect obviously lies in the double
doses of compensators for these normal genes which the female possesses. And that the two
doses in the male cause even more abnormality than three doses of the same section of
chromosome in the female is also to be expected, if the degree of effect depends on the
ratio of primary genes to compensators. For the two-dose male has a ratio of these (2:1)
which is further from the normal ratio (2:2 or 1:1) than is that of the three-dose female
(3:2). |
|
All the above evidence shows
clearly that the normal genes in the X-chromosome are subject to dosage
compensation, even though we can seldom prove this directly in the case of previously
selected loci, containing the normal alleles of known mutant genes, since most normal genes are too near their saturation level to make such
observation possible. But the question then arises, should not the very fact that most
of these genes are so near their saturation level make dosage compensation unnecessary in
their case? We have seen
that for the normal alleles of apricot, scute, etc., practically no difference between the
effects of one and two doses can be seen in either female or male. Why then would there be
a perceptible advantage in going through the motions of equalizing them still further? The
answer can only be that since, even for these cases, there is evidence from the mutant
alleles that the compensation mechanism does apply to the given locus, we must conclude
that there is an advantage in it, even when the genes are normal ones. |
|
[Commentary:
In this delicious section, Muller, with impeccable logic, begins to dig himself into the
metaphysical hole referred to above. "Manifestations
of dosage compensation are all
exhibited by mutant genes (mainly hypomorphs)", because they "have a
level of activity so low that differences in their dosages or activity give an easily observable result, whereas for normal genes, lying as they do near their saturation
level, the difference in expression resulting from two as compared with one or three doses
are usually imperceptible". But mutant genes
occur so rarely in the population that they are unlikely to be the target of natural
selection. The dosage compensation "effect on
the manifestation of the mutant character must therefore be an automatic result of the
operation of processes" which have nothing
to do with the characters themselves ("other
connection").
Muller infers a general
over-riding mechanism, of unknown role, ensuring that, for the majority of
genes on the X-chromosome, "the
grade of development of their effects, which is best for one sex, is pretty much the same
as the grade for the other sex".
(Only in the case of the few genes concerned with sexual
differentiation would there be "some
advantage to the individual and the species" in having sexually distinct levels of expression.) Consistent with
an over-riding mechanism, otherwise non-mutant females, aneuploid for part of one X
chromosome, show decreased viability, even though they now have the same dosage of the
genes corresponding to that part as the male. This appears as a collective function "so
that, by the reduction to single dose of many at once, the combined effect becomes perceptible, where the individual
effect would not be".
For normal genes, there is "practically no difference between the effects of
one and two doses … in either female or male". So, "why
should there be a perceptible advantage in going through the motions of equalizing them
still further? …We must conclude that there is an advantage … even when the genes are normal
ones". What could that evolutionary advantage be? The possible
importance of the collective function is stressed elsewhere (Forsdyke 1994; below)] |
THE SURPASSING PRECISION
OF ADJUSTMENT THEREBY IMPLIED
Let us now try to estimate how
small the sex difference in the character
is, which we have thus decided must affect the organism's welfare. We may use here
essentially the same method as we applied on pp. 182-3 when we calculated, on the basis of
the variability of the mutant, how much variation in the normal character was consistent
with survival. We may assume, as a first approximation (though one justified by existing
data on multiple alleles), that the amount of effect of the compensators on the mutant
genes is ordinarily about the same, relatively to the activity of these genes, as the
amount of effect on the normal genes, relatively to the activity of the latter. That is,
since we observe that the additional dose of compensators present in the mutant female as
compared with the male, reduces the effectiveness of the two doses of the given mutant
gene which she possesses so as to make them equal to the one dose of the male, we infer
that the compensators are likewise, in the normal female, reducing the effectiveness of
her two doses of the normal allele to that of the male's one dose, and with at least as
great relative accuracy. We may tentatively mark off the amount of activity thereby
attained in the normal female as a point on the abscissa of our curve (e.g. in Fig. 5),
placed in a region so near that of the saturation level that a point half way from the
origin to this point would still give a character not perceptibly (to our unaided
observation) different from it.
Now there has
necessarily been a certain latitude in the determination of the exact grade of
effectiveness of the mutant allele (e.g., apricot), owing to:
Moreover, in the case of some mutant
genes, there is also some inaccuracy in the compensation achieved, evinced by under- or
over-compensation as the case may be. A maximum estimate of the error from all these
causes together has been indicated, in the case of apricot, by the dotted vertical lines
arising on either side of the ordinate that marks the typical effect of two doses of the
mutant in the female or one in the male. (In the given case, these limits represent mainly
the degree of coarseness of our powers of observation -- i.e., the discrimination
thresholds.)
But, in
harmony with our previous premise, we should, to be on the side of caution, assume that,
in its action on the normal character, the process of dosage compensation has an
equivalent amount of latitude to that found in the mutant, in the sense that the
inexactitude may be as much, relatively to
the total activity of the normal genes, as it is relatively to that of the mutant genes.
This enables us to plot on the abscissa the approximate plus and minus limits of the
effectiveness of the genes existing in the normal type (either male or female).
Constructing ordinates at these limits, and marking off the points at which they intersect
our curve of effect, we may then find (on our vertical axis) the maximum difference in the
normal character which could be produced by one dose in the normal male as compared with
two doses in the normal female. It will he seen that in the given case this difference,
which might be called the maximum inaccuracy of the dosage compensation, is small, even as
compared with the difference caused by a change from one to two doses in the same sex.
Stating the above in
a somewhat different way, let us first assume a maximal inexactitude of compensation for
the mutant gene studied (apricot). This would here be about equal to one step of visual
discrimination, since (according to observations of the author but contrary to some
statements in the literature which report the male to be slightly darker) no certain
difference can be distinguished between the apricot male and female. Now divide this
maximal inexactitude (or, more properly speaking, uncertainty) by the amount of visible
difference which the compensation mechanism is here working to offset, i.e., by the amount
of visible difference between the effects of one and two doses of the apricot gene in the
male.
This gives a
relative value for this maximal possible inexactitude, which turns out to be about one
tenth. That is, at least nine tenths
of the dosage difference has been compensated for, but possibly (at most) there is a one
tenth under- or over-compensation. To be on the side of caution, we may now assume that
there is as great a relative inexactitude of compensation for the normal alleles also --
although, on the average, the inexactitude should actually be less for normal than for
mutant alleles, on account of the mechanism having been evolved in relation to the
normals. This then leads us to the conclusion that, in the case of the normal alleles of
apricot, not more than one tenth of the difference between the effects of one and two
doses in the male has escaped compensation, while nine tenths at least have been
compensated for. |
|
But
we have seen that the entire difference in question, between the effects of one and two
doses of the normal gene, whether in
male or female, is itself (on the average, at least) imperceptible to our eye. Thus we find that selection has been sensitive to differences at
least an order of magnitude smaller than the smallest difference which we can ordinarily
see. Hence even subliminal gradations of this fineness are actually of adaptive
significance to the organism.
|
Application
of the same method leads to similar conclusions in the case of scute and most of the other
characters observed in regard to this point. That is, the compensation mechanism involves
a process, or system of processes, that works to a much greater degree of refinement than
is visible to us. We have seen, then, that natural
selection has in the first place, in the interests of character stability, established the
effectiveness of the normal gene at a point on the curve so near to the saturation level
that one dose is not visibly different in effect from two, so far as
we can see. Yet even this likeness is far from sufficient, for organisms (male versus female)
that may differ regularly to this seemingly slight degree, otherwise the mechanism of
compensation would not have been set up, in addition to this, in the case of the great
majority of sex-linked genes, so as to reduce the difference to a tenth or less of even
this invisibly small amount!
It is evident too
that the required equalization cannot so advantageously be achieved by the simpler method
of stepping up the normal gene's effectiveness still more, into a still flatter portion of
the curve, otherwise the more elaborate compensation mechanism which exists would not have
been needed. That it does exist, and therefore was needed, shows that a relatively slight
increase in the effect, in fact imperceptible to us, brought about in the above way, is
disadvantageous, just as an imperceptible decrease in the effect is.
In other
words, it is as important to keep down the grade of the normal character to the given
level in the two-dose female as to keep it up to that level in the one-dose male, and in
most cases the gene's activity cannot advantageously be brought near enough to an actual
saturation level to obviate this requirement. However, for the lesser variations in
effectiveness of the normal gene, caused by fluctuating environmental conditions and by
variable genetic "background", the raised stability
brought about by the near-saturation level of the gene must be of great value.
There is no doubt,
however, that in many cases specific regulatory mechanisms
("servomechanisms") have been evolved, in addition to the
above, to achieve an even higher stability in the face of given especially frequent or especially disturbing conditions,
as many phenomena both of embryology, regeneration, physiology and pathology show. Moreover, in a relatively small but absolutely huge number of
cases, the adaptive reactions go far beyond mere stability, and consist instead of
positive changes of an advantageous nature following given types of changes in conditions.
These reactions range from the "morphogenic" to the
"physiological," there being no essential
distinction between the two categories. This however is a topic too large to be entered
upon here, since it would launch us upon the entire subject of how the organism works. We
mention it here only to set our own problem in its proper limited place in relation to
biological phenomena in general.
In order to avoid the
conclusion that differences far too small to be evident to us are nevertheless of
importance to the organism, some might prefer to suppose that selection has rather worked
by eliminating the occasional individual in which, owing to special conditions, the
character in question had been influenced to a considerably larger degree than usual by
the gene difference in question. This possibility has already been discussed in its
relation to the character stabilization brought about by raising the gene's effectiveness
to near-saturation levels.
We have seen
that, although this position is tenable, and is doubtless valid to a certain extent, it
nevertheless brings us to much the same conclusion in the end, when this is expressed in
terms of the average amount of
character-difference caused by a gene difference which has an appreciable adaptive value.
Moreover, there is reason to infer that, in many cases, there is no actual threshold
amount of difference which suddenly emerges as disadvantageous, but that the amount (or
the chance) of disadvantage may be taken as roughly proportional to the amount of
deviation from the norm, even in the case of minute differences.
In such
cases, then, the amount of selection involved would be the sum of that occurring for each
grade of expression, and that for a given grade would be proportional to approximately the
product of the number of individuals with that grade times the amount of their deviation
(i.e., the size of the grade, when the normal is designated as 0). A reckoning of this
kind will show that here the selection based on relatively low grades of deviation (e.g.
those less than the standard deviation) will probably in most cases considerably outweigh
that based on the rarer, more extreme deviants. This presupposes, however, for the cases
we have dealt with above, that survival affords a considerably more sensitive criterion of
the degree of development of the character than does our own eyesight. |
|
[Commentary:
Muller here proposes that: "in its action on the
normal character, the process of dosage compensation has an equivalent amount of latitude
to that found in the mutant". Then, he proceeds
to infer "that selection has been sensitive to
differences at least an order of magnitude smaller than the smallest differences which we
can ordinarily see.
Hence, even subliminal gradations of this fineness are actually of adaptive significance to the organism". Having noted in previous sections that:
-
(i) dosage compensation reflects processes
having an affect "in some other connection", and
-
(ii) to detect the influence of dosage
compensation with normal genes requires an experimental situation where many of them are
affected collectively,
Muller now resorts to explaining the
evolution of dosage compensation in terms of Natural Selection acting "in the interests of character stability" on exceedingly minute individual gene character
differences, "otherwise
the mechanism of compensation would not have been set up". Furthermore, selection "based on relatively low grades of deviation …
will probably in most cases considerably outweigh that based on the rarer, more extreme
[specific character] deviants".
However, he
remains aware of the possibility of some
more general selective process "evolved, in addition to the above, to achieve an
even higher stability in the face of given especially frequent or especially disturbing conditions".
Among these we might include the heat-shock response (Forsdyke 1994).] |
AN
OBJECTION BASED ON CORRELATION BETWEEN USELESS AND USEFUL CHARACTERS
A different objection which may be raised to these conclusions is
that, in studying the above characters of eye color, bristle
development, etc., we may have been dealing only with certain unimportant
superficial expressions of gene changes which have other, more important, effects,
hidden from our casual observation. In other words, characters are often correlated in their development, and we may have studied
the less important by-products of the gene-initiated processes.
On this view, natural selection may in truth have resulted in the establishment of
dosage compensation for the normal gene, because of the advantage of attaining a given
level of development of its most important effect. This effect
itself, though hidden, might be subject to a very considerable dosage influence. Yet,
studying only the side effect, we may have been able to obtain evidence of only a
subliminal action of the compensators. Granting this
viewpoint, we might avoid the conclusion that selection has had to deal with extremely
fine differences.
It is true that genes
do have multiple effects, thus causing correlations between characters, and that in no
individual case with which we are dealing can we know that the advantage of the dosage
compensation mechanism lies in its influence over that particular effect of the given gene
which is obvious to us. However, it would be strange if we
had missed the major effect of the gene in such a large majority of the cases,
especially since some at least of the genes concerned (the normal alleles of yellow,
achaete and white) have been proved physiologically dispensable, i.e., the individual can
live in the total absence of these genes. But, if we have caught the major effect in any
of the cases, then that effect does show the type of compensation at issue -- a type so
refined as to operate on subliminal grades of the character. Furthermore, we have seen
that with but very rare exceptions (relative to the total number of genes) the effects of individual genes, whether in the X or other
chromosomes, are so near their saturation level as to make direct discrimination between
one and two doses impossible. Since a very wide sampling has consistently
given this result, it would be highly unlikely that this relation
did not hold for the major effects of genes in general as well as for their minor effects.
In fact, if our
interpretation is correct it is particularly for the major effects that this aid to
stability has been established. But if this is true of the major effects, and if the
dosage compensation found is, as the present criticism started out by assuming, also a
phenomenon developed primarily for its influence on the major effect, then the main
conclusion still remains valid, for the compensation results in much finer grades of
difference even than those, already subliminal, which are due to doubling or halving the
dose. Thus, in any case, the compensation mechanism must be
concerned with the equalization of exceedingly minute differences.
A third point to be
considered in connection with the question here at issue is the following: The great
majority of characters which a gene affects are not immediate products of it but are
end-results of complex webs of biochemical processes, in which a given thread or chain of
reactions can, theoretically, be picked out that has its root in the gene in question, and
leads from the latter to the character through a considerable series of events (see Fig.
7).
Now when
the given gene has multiple effects, it is evident that the gene either serves as the root
of two or more entirely separate chains of reactions which respectively lead to the two or
more characters affected, or else that, although but one basal chain of processes has its
start at the given gene, this chain somewhere along the line becomes branched, so as
finally to diverge to the two or more correlated characters. Owing to the complexity of
the whole web of ontogenetic and physiological processes, the branching will, in the case
of different genes, occur at different points. In some cases it will happen to occur
early, near to or at the gene itself, while in others it will occur late, near the
character which the observer examines. This diversity in the position of the branching has
been amply demonstrated by many studies in physiological genetics, as well as in other
fields of biology and medicine. |
|
|
Now influences derived from other genes (as well as, ultimately at least, from
environmental sources) impinge upon these chains of reaction at practically every link,
thus playing their part in the determination of the final characters. Unless we make very unusual assumptions the compensating genes work in
this way too. That is, the compensations may be achieved by influences impinging on
the reaction chain at any point. |
| FIG. 7.
Diagram of a branching chain of reactions whereby a given gene produces two effects. Each
straight continuous line represents some intermediate process between the gene and the
characters observed, and each dot represents a new developmental stage or substance,
resulting from those processes which lead up to it. Dotted lines represent processes
derived from other genes or gene combinations, or from environmental influences, that
impinge upon the processes derived from the given gene and affect the nature of the next
stage reached. |
And as it is a matter of indifference to the organism
at which point this anastomosis of effects takes place, so long as the final advantageous
result, compensation, is attained, it will sometimes be above and sometimes below the
branching that leads to a given character (Fig. 7), if this character alone is important
enough to have compensation established for it.
Moreover,
in such a case, if the compensation is the accumulated result of a number of influences,
derived from different, cooperating compensator-genes, some of these influences,
ordinarily, will be brought to bear above, and others below, the point of branching. Only
those influences which act below the branching, however, would then affect the secondary
characters as well as the important one. For this reason, we should not expect to find
dosage compensation to be nearly so widespread, or so accurate, as it is if we had been
studying only secondary characters, such as were of too little importance for the
mechanism to have been developed in relation to them themselves. |
All in all, then, we may reject the objection based on correlation of characters, and conclude that dosage
compensation has in fact become established because of its advantage in regulating more
precisely the grade of characters whose variation in grade, even without it, would
be exceedingly minute. |
We may
here digress to point out that the considerations above set forth concerning the way in
which the reactions initiated by genes bifurcate to produce multiple effects (Fig. 7) also
throw light on the related question of the extent to which selection for a given important
character may be expected automatically to carry, in its train, a developmentally
correlated but not advantageous change in another character.
It should
rarely happen that two apparently separate characters, especially if they be characters
governing only degrees of development or proportions of already present parts or
substances, will be so closely connected, with the branching occurring so very near to the
distal end of the reaction chain of either of them, that they cannot both be affected
separately, by influences impinging beyond the point of branching.
Therefore, if the so-called secondary effect is a disadvantageous one, it will usually be
corrected, without disturbance of the primary effect, by means of a selection acting upon
genes which affect the secondary branch but not the main stem. Moreover, it is to be
expected -- especially in view of our present results -- that most visible characters,
even those of an apparently superficial nature or of very minor degree, will nevertheless
have enough importance in themselves for their variations to be felt in the selective
process. For this reason we should be very wary of any interpretations which would explain
the existence of a given grade of development of a character as an automatic by-product of
selection for a more primary character.
So, for example, the
finding that the mutation from the normal grayish-tan to yellow body color in Drosophila
results in a cuticle which is not so resistant to environmental changes in dampness, in no
way explains why the flies do not normally have a yellow body color. For if that color
were in itself advantageous it could doubtless be arrived at by selection of mutations in
other genes which, though not having the undesirable chemical action, nevertheless gave
effectively the same color, or by selection of "modifying"
mutations which, with the gene for yellow in question, counteracted the undesirable
chemical effect, while leaving the color yellow. In fact, there are other species which
normally look rather like the yellow mutant of D. melanogaster and it is probable
that in them some such arrangement has in fact been attained by selection (without giving
them the above mentioned handicap), as a result of ecological conditions which put a
premium on such a color.
The organism, then,
is genetically highly plastic, at least so far as its proportions and the relative degrees
of development of its different features are concerned. Thus we may assume it, in general,
to have attained optima in all these respects, largely regardless of developmental
correlations. In fact, the correlations normally occurring, such as the observed
principles of heterogonic growth, Gloger's rule, etc., have themselves come down through
the long mill of selection and hence are themselves in large measure advantageous adaptive
reactions. Moreover, they can themselves be changed again by further mutation and
selection, as observations of specialists in such fields have shown. Nevertheless, the
organism does of course have its grave limitations, causing change in some directions, and
to some degrees, to be more difficult than in others, or even impossible. |
|
[Commentary:
Darwin and Wallace both used the unknown-but-correlated-character argument
when they were pressed for an explanation regarding utility of a character with respect to
the action of Natural Selection. Muller explores this here, showing that he is firmly wedded
to explanation based on gene-specific characters, whether correlated or
non-correlated.]
|
THE DEVELOPMENTAL
MECHANISMS INVOLVED IN DOSAGE COMPENSATION
In the above discussions we have implied that dosage compensation
has, for each sex-linked gene that is subject to it, become established through a natural
selection of all available and innocuous mutations in the X-chromosome that chanced to
result in a compensating action upon the expression of the sex-linked gene in question.
(Included here of course would also be such mutations in the given gene itself as
furthered its sensitivity to such compensation.) This interpretation would put the method
of evolution of this phenomenon in line with that which we have good reason to conclude
has been followed in the case of most other functional systems of organisms.
It might
however be postulated that there are one or more peculiar genes in the X-chromosome
which are especially suited to serve as compensators to all others in it, and that the
only mutations then needed to achieve compensation are those, perhaps of a simple specific
type, in the genes to be compensated, which allow the developmental processes caused by
them to become responsive to the influence of these particular compensators. As a
plausible guess, it might further be postulated that these compensators are none other
than the gene or genes determining sex itself, and that their compensating influence is
exerted through processes which form a part of those that differentiate the sexes.
Even though it is now
known that, in Drosophila, the turning of the scales which decides
which sex is to develop depends upon the cumulative action of numerous
sex-differentiating genes of individually small effect, scattered through
the X-chromosomes, this would not vitiate the possibility here in
question. For the effects of these numerous genes obviously converge to set going one
fundamental sex-differentiating process, or "focal reaction",
on which in turn most or all of the secondary features of sex differentiation depend (see
Muller, 1932a).
This is shown,
among other things, by the essential similarity of the whole system of results, so far as
sex is concerned, whenever any limited group of these sex-differentiating genes comes into
play, in experiments in which the dosage of different parts of the X-chromosome
is varied. Thus it might well be supposed that either this main sex-differentiating
process itself, or one derived from it, served as the root mechanism for dosage
compensation, which would thereby take on the aspect of a secondary sexual phenomenon of
fairly familiar type.
This possibility was
never thought to be very probable, in view of certain results derived from a study of X-chromosome
fragments, to be mentioned presently. It has, however, been pretty definitely disproved
recently by some experiments carried out in 1946 by the present writer and Miss Margaret
Lieb in collaboration, but not yet published (see Lieb, '46). These made use of a mutant
gene called "transformer" found by Sturtevant, a
recessive lying not in the X but in the third chromosome, which has the
effect, when homozygous, of causing flies with two X-chromosomes, which
otherwise would have become normal females, to develop into males.
Although
sterile (even when provided with a Y), these males are well developed in
virtually all other respects, so that it can be inferred that the main sex-differentiating
process has been initiated in them in very much the same form as it has in regular males.
If now the dosage compensation of the Drosophila female is brought about through effects
of the female sex-determining process, or system of processes, as such, then these males
with two X-chromosomes should not have the dosage compensation characteristic of the
female operative in them. If, for instance, they are provided with two doses of apricot,
one in each X-chromosome, they should have the rather dark eye color characteristic of the
male with two doses (as in the rightmost eye of Fig. 4), not the compensated color of the
female with two doses, which looks like the one-dose male.
The test
clearly showed, however, that they had the fully compensated color, like that of the
female with two doses, even though they were males with two doses. Obviously
then, the compensation was caused, as in the ordinary female with two doses of apricot, by
the presence of a double dose of compensators, lying in other parts of
the X than at the locus of apricot itself, and these worked quite independently of
the sex-differentiating genes, since all of the latter, together, although
present, were here quite incompetent to cause any visible approach towards the female sex.
Tests of several other compensated sex-linked characters than apricot -- scute, forked and
Bar -- with the transformer gene, gave this same result quite unequivocally for all.
These findings had
been expected because tests performed much earlier, involving the addition of different
fragments of the X-chromosome, first by the writer and then by Offermann
working in collaboration with the writer, had shown that, in the case of different
sex-linked genes, there is a different distribution of sex-compensating potencies
throughout the length of the X-chromosome. That is, not only were the
fragments taken from different regions unequally effective in the compensation of a given
gene (when due allowance was made for their sizes), but their effectiveness relative to
one another was not the same when tested with one sex-linked gene as when tested with
another one. The different genes therefore had, to a considerable
extent at least, different compensators, not one common set of compensators as they would
have had if the compensation had worked through sex-differentiating effects.
Furthermore, the results for none of the sex-linked genes agreed well with the relative
values of the different regions of the X-chromosome in influencing
sex-determination itself, as worked out, after some pioneer work of the present author,
chiefly by Dobzhansky and Schultz, and by Patterson, Stone, Bedichek and their associates.
A further and more
elaborate investigation of the behavior of the different regions of the X-chromosome
in compensating for particular genes has been carried out more recently (1946), by the
writer and Lieb working in collaboration, and the results from this confirm and extend the
earlier conclusions. It would takes us too far afield here to describe in detail the
technique or the results of these various experiments on the location of the compensators.
The method was, essentially, first to produce by irradiation fragments of the X-chromosome
taken from different regions, and then to determine to what extent the addition of these
fragments to a regular chromosome complement (usually of a female) affected the expression
of a given sex-linked character, the locus of which lay outside of the fragment.
In most cases
the compensation proved to be a complex phenomenon, which could not be pinned down, in
toto, to a particular chromosome region, nor even regarded as a simple summation of
the effects of the different regions. Some regions, however, had distinctly more influence
than others in the compensation of a given gene, and there were even cases where a certain
region exerted what may be called a "negative compensation",
working in exactly the wrong direction, an effect which was however counteracted by means
of overcompensation elsewhere. That is, the surprisingly accurate compensation exerted by
the chromosome as a whole was a final resultant, brought about only through an integration
of individual effects which in detail appeared chaotic. And a similar chaos was evident on
comparison of the manner of distribution of the effects among different regions, as seen
in comparisons of the cases of different compensated genes.
Now this is exactly
the sort of result we should anticipate if each gene has had its own
separate system of compensators selected for it, and if the compensation for each
has, in most cases, come about by a succession of steps. Some genes could of course have
gained a foothold (if we suppose them not to have been priorly present) that even worked
the wrong way in the given respect, provided they possessed an advantage lying in some
other direction, but even these would ultimately be corrected, by the selection of genes
that counteracted such an effect, as we have pointed out in our discussion of correlated
characters, and so the final resultant effect would be superior to that of any single
interacting gene. The attainment of the accuracy finally arrived at must usually have
required many small mutations, which, taken together, increasingly whittled the character
into shape, as it were. And while in some cases these mutations may have been successive
changes in the same gene, any mutation in the X-chromosome that worked in
the advantageous direction would be acceptable, so that in most cases it is to be expected
that a fairly complex system became at last established. Since,
however, the advantage of the compensation for any single locus is only what might be
called a "second-order" one, dealing as it does with such minute, to us
subliminal, grades of effect, the establishment of these precisely working systems must
have taken far more time than needed for visible "first order" effects.
Moreover, there may more often have been setbacks which required repair, and which thus
led to still greater complexity.
If we may infer that
most of the genes in the X-chromosome have compensation systems like
those studied above (an inference to be accepted with some caution since we were dealing
only with genes capable of giving conspicuous visible mutations), we would be led to the
peculiar conclusion that, since there is usually more than one compensator for each gene
of the X-chromosome, and since the compensators are themselves in this
chromosome, most sex-linked genes not only have an original or primary
function but also serve as compensators for one or more other sex-linked
genes. Each individual gene then would be complex in its functioning and also in
its potentialities of mutating so as to affect its different functions to
different degrees, relatively to one another. Finally, as there are over a
thousand sex-linked genes, it will be seen how amazingly complex, in all probability, is
the tangle formed by the sex-linked system as a whole.
In conceiving the
mode of action of a compensator, or collective group of
compensators, for a given sex-linked gene, which we will call the
"primary gene," it is easy to make the mistake of thinking that each dose of the
compensator or compensators effects a given total amount of reduction of the activity of
the primary gene (or rather, gene-product). If this were true, however, then the female
with her two doses of both primary and compensators would show an effect equivalent to
twice that produced by the one dose of primary and one of compensators in the male. In
other words, there would be no equalization of the sexes. Moreover, in that case
compensators acting strongly with hypomorphs would not work properly for genes at higher
levels. We must therefore infer that the compensators, when present in any given dose,
work in such a way as to effect the same proportionate amount of reduction in
primary gene activity, regardless (within wide limits) of what the dose or activity of the
primary gene is. This would ordinarily be the case if the inhibiting action of the
compensator were itself little influenced, in return, by the amount of primary
gene-product it had to affect. An example of this would be a
situation in which the compensator's "product", determined indirectly by its
gene, consisted in some such pervasive condition as a relatively high pH,
which the primary gene's product, no matter how concentrated, had little effect on. In
such cases, then, the compensators, at a given dose, would tend to reduce the primary
action by a given proportion, rather than by a given absolute amount.
Moreover, the
relations must be so fixed that the compensators, when themselves in double dose, reduce
the effect of a given dose of the primary gene to half of that which would obtain in the
presence of a single dose of compensators. For only thus can the effect of the female's
double dose be reduced to that of the male's single dose. Such a result would be brought
about most simply in a case in which the compensators, when themselves in single dose,
reduced the primary effect to half what it otherwise would be, and in which, when their
dose was raised, their own effectiveness rose in the usual geometrical manner. For, in
such a case, one dose of the primary with one of compensators, as in the male, would have
an effect which may be designated as "one half" (i.e. one half that of one
uncompensated dose of the primary gene). And two doses of the primary with two of
compensators, as in the female, would have an effect equal to the product of 2 doses (for
the primary) times 1/2 (for the first dose of compensators) times another 1/2 (for the
second dose of compensators), a product which of course is itself equal to one half, and
therefore the same as the effect in the male, i.e., properly compensated. And no matter
what the degree of activity of the primary gene might be in such a case (e.g., whether it
were the normal allele or, say, a hypomorph having an activity equal only to one quarter
of one dose of the normal), the compensation would still work to equalize the effects of
the one and two doses of the primary in the two sexes. Whether this simple scheme is
usually true can probably be determined definitely through quantitative studies involving
several different doses of compensators. |
|
Commentary.
Here Muller struggles to understand how the postulated compensator genes would operate,
resulting in vague but interesting suggestions that "sex-linked genes not only have an original or primary function,
but also serve as compensators for one or more other sex-linked genes. Each individual
gene would then be complex in its functioning and also in its potentialities of mutating
so as to affect its different functions to different degrees...".
There might be a "collective group of compensators" with a "product",
determined indirectly by genes, consisting of "some such pervasive condition as a relatively high pH", which "would tend to reduce the primary action [of a given gene] by a given proportion, rather than a given absolute amount". However, one may comment that it is difficult to
see how such a "pervasive
condition" would avoid affecting
also the products of autosomal genes. |
EVIDENCE
OF THE SECOND-ORDER PRESSURE OF SELECTION IN ESTABLISHING DOSAGE COMPENSATION
It has been mentioned that, according to the conception above
outlined, dosage compensation must be a "second-order"
evolutionary phenomenon, since the selective force that establishes it must depend upon
such minute advantages. It might well be so slow, therefore,
as not to have caught up with the sex-linked genes in species in which they had been
established as sex-linked for a period of the order of only a few million years.
There are several known Drosophila species which probably meet this condition, and which
might therefore serve as a partial test of our thesis. Of these, the most worked with has
been Drosophila pseudo-obscura, and the results with it have therefore been
reexamined from this point of view by Dr. Rowena Lamy and the present writer, in
collaboration.
A considerable body
of earlier work had joined to show that what serves as the X-chromosome
in D. pseudo-obscura consists of two parts or arms, one of which is, for the most
part at least, descended from the original rod-shaped X-chromosome, that
has remained as the X also in our common species melanogaster, while
the other arm is, as a part of an X, a relatively new acquisition, since
it corresponds to what forms the left arm of the third chromosome of melanogaster, and
also lies apart from the X system in most other Drosophilas, as well as,
presumably, in the common ancestor of both pseudo-obscura and melanogaster. The
connection of it with the original rod-shaped X, to form a two-armed X,
must have occurred by what we call a whole-arm translocation, and this must have been in a
period geologically a good deal more recent than that at which most of the material of the
original rod X was established as X-chromosome material.
In view of this, it might well be expected, on our conception, that while the genes of the
original X arm would show good dosage compensation, as they do in melanogaster,
those in the other arm might still have this mechanism much less perfectly evolved.
For testing this
question, we have not resorted to the difficult and rigorous method of studying dosage
effects as revealed by the addition or subtraction of fragments. We have, however, made
use of the knowledge, derived from melanogaster, that many or most mutant genes are
hypomorplic and, unless compensated, exert a more nearly normal effect in double than in
single dose. This being the case, most of the mutants in a well compensated part of an X-chromosome
would look alike in male and female, whereas many of those in a poorly compensated part
would look more extreme in the male. There would, to be sure, be exceptions caused by the
fact that some mutants are partially "sex-limited"
or "sex-influenced". This phenomenon, which is most
readily demonstrated in studies of non-sex-linked genes, but which must apply to some
sex-linked genes as well, is quite apart from dosage compensation, and probably represents
in some cases an adaptation to differing sex needs. However, the deviations from equality
of sex-expression which were produced in this way would be just as likely to cause the
male to be less extreme than the female, as to cause him to be more extreme, and they
would not have a tendency to affect genes in one chromosome arm rather than in another.
When the results of
sex comparisons were tabulated for all those sex-linked genes concerning which data were
available to us, it was found that in the "left"
arm of the X of pseudo-obscura the great majority of the mutants looked alike in
the two sexes, with however an occasional exception, that in one case had
the male more extreme, but in the other case, the female. Now this is the arm that is descended from the original X-chromosome.
In the more recently acquired arm of the X, on the
other hand, the "right" arm, there was a much higher proportion of genes which
showed a sex difference in their grade of expression. Moreover, in nearly all these cases
the direction of the difference was the expected one, namely, that in which the female
stands nearer to the normal type -- an indication of absence or incompleteness of
compensation.
As yet, the numbers
of mutants examined in the above way are hardly great enough to make our conclusions as
secure statistically as we would wish, yet the agreement with the
theory is fully as good as we had expected. This evidence can therefore be counted
as contributory to our general thesis, especially since it comes from such a different
direction of attack than the previous work. It would be of great interest to have further
evidence, involving exact dosage studies, obtained along this line. This would be
especially valuable in such a species as Drosophila miranda, which is closely
related to D. pseudo-obscura, but has been found by Dobzhansky and Tan to have, in
addition, a third portion of the X-chromosome system, still more recently
acquired by the latter. Drosophila americana and virilis would likewise provide an
important comparison for this purpose. |
|
Commentary:
Genes recently translocated to the X-chromosome do not
show dosage compensation. For Drosophila, this implies a local
compensating mechanism, rather than a global chromosome-wide
mechanism.
|
EVIDENCE
OF ADAPTIVE PRECISION BASED ON THE PROLONGED PERSISTENCE OF MINUTE FEATURES
Although the facts concerning dosage compensation appear to give
us the most searching evidence of the amazing degree of precision of genetic adaptation,
or, to put the matter conversely, of the disadvantageousness of even very subliminal (to
us) departures from the already established type, nevertheless we have not to look far to
find other lines of evidence, of somewhat lesser exactitude, for the same sort of thing.
Among these, one of the most convincing, from a genetic standpoint, is the very slow rate at which most characters of most species are, at any given
moment, undergoing evolutionary change, as shown by the very high resemblance of the
organisms in most respects to their ancestors of, say, a million years before.
True, there
have been other periods, of comparatively rapid evolution, for each line of descent, as
for example in the human stock during the past few million years, but these are relatively
rare. They illustrate the general principle, applicable in biological evolution as
elsewhere, that "one good turn deserves another".
On the whole, during the past of most species, the periods of high stability have occupied
much the greater portion of the duration of their lineage. Thus many insects preserved in
the Baltic amber were, in the pettiest details, sensibly the same species at the remote
time when they became thus entangled as they are today.
That this stability is due to active selection in favor of the normal type,
that is, to the fact that the latter is more advantageous than even the slight deviates
from it, is shown by a consideration of the findings of genetics concerning the frequency
of mutations. In those few but very different multicellular organisms which have served as
samples for the investigation of this subject, the frequency of detectable mutations in
individual genes has proved to have a range of values centering around the order of
magnitude of one such mutation in the given gene in about fifty or a hundred thousand, or,
at most, a few hundred thousand, germ cells. Or, if we were to follow a given gene
down through a single line of descendants, this would mean that it would on the average
undergo such a mutation in fifty to a few hundred thousand generations. In most higher
organisms, this would occupy less than a million years. If slighter mutations than those
easily detectable could have been taken into consideration, the period between one
mutation and another of any given gene which we were following might in fact have been
considerably less than this. Now if the mutant genes did not
handicap the individuals in their living, but had an equal chance of survival with the
normal, it would in consequence come to pass that after a million years had elapsed the
great majority of genes, and with them practically all characters, would have become
appreciably different. And in just a few million years the organisms would have become
practically unrecognizable in all respects, merely as a result of "mutation
pressure".
We may, to be sure,
grant that many mutations are so pronounced or so manifold in their effects as to be
lethal or detrimental, and that a change in all genes would therefore be too pronounced a
result to expect in the above period. However, despite this stricture, it remains to be
taken into account that practically all characters depend on many
genes, and that of these a large proportion produce only minor effects on them [the
characters] and often mutate in such a way as not seriously to affect other characters at
the same time. Therefore, if we considered the mutation frequency
for a given character instead of for a given gene, we would find it to be very much
higher than the above, even when only those mutations were included in the reckoning which
caused but slight changes in the character and which produced little in the way of
detrimental effects of other kinds. As a consequence, if these slight changes in the
character were not in themselves disadvantageous, a few million years could not
pass without practically every character having undergone a whole series of stepwise
changes, more or less random in their directions, some steps having been smaller,
some larger, and all combined having the result of making the character very different
from what it had been. The only preventive to this unsettling effect of the prolonged
mutation pressure would be selection, -- i.e., the dying out of
the mutant individuals along the way, -- by reason of the disadvantage conferred by the
slight, as well as by the great, variations in the character. Thus, the mere presence of a given character, or of a given grade of development
of a character, in a species, over a period as long as a million generations, in itself
demonstrates that this character, or even this particular grade of development of it, is
of material value to the organism in its struggle for life and perpetuation.
It is evident that
this line of argument becomes even stronger in those numerous cases
in which a given character, or a given grade of its development, is found in two or more
related species, that probably diverged from a mutual ancestral type some million or more
generations previously. In such cases no records of this ancestor itself are
needed, provided there is good ground for it to be at least as remote as this. But there
is sometimes a further kind of argument in these cases. This is illustrated by
Sturtevant's finding of many years ago, that when the related
species Drosophila melanogaster and D. simulans are crossed, although both
these species have an identical pattern of bristles, one which must have been very long
established and present in their common ancestor, nevertheless the hybrids between them
show considerable disturbance in this pattern, and especially a tendency to have various
bristles missing.
This can only mean that the present developmental mechanism of bristle production
in one species has come to have such a different genetic basis from that in the other
species that a mixture or compromise between the two system involves incompatibilities
which obstruct the attainment of the effects of either system alone. It is thereby proved
that, while the given character remained in, or regained, its original form during the
evolution of both species, nevertheless the genetic and developmental basis beneath it
underwent changes.* [Footnote: Better instances are provided by cases
in which the introduction of a particular gene (mutant or normal) for a given character
from one species into another by backcrossing (or, as Edgar Anderson terms it,
"introgression") causes a change in the expression of that character, even
though the normals of the two species had looked much alike in the given respect. Such
cases have been reported by Harland in cotton, by Gordon and by Kosswig in fish, by the
present writer in Drosophila, etc. When the conclusions are based only on the first
generation hybrid it might be supposed that incompatibilities involving other genes, not
normally affecting the given character, were causing disturbances that now affected it.]
Surely the above
changes would not have been just so adjusted as to allow the development of a character
which was outwardly the same as before unless there had been some controlling agency which
allowed only such changes to become established as failed to alter the end-result (here,
the bristle pattern), or which allowed only such changes as were effectively counteracted
by others which maintained that end-result. Such control, in the interests of the
end-result itself, rather than of the mechanism behind it, can only signify (if we reject
the animistic interpretation of a conscious guidance) an active selection favoring
individuals that exhibit this end-result.
In other words,
the advantageousness of just this result for the organism is thereby implied. The numerous interspecific crosses in which, even though the
first-generation hybrids may appear normal, some of the types produced by genetic
recombination in later generations nevertheless manifest abnormalities, show equally well,
though slightly less directly, that here too the genetic basis had become different, while
maintaining a similar facade. These cases then greatly extend the number
of examples of characters that must have been forcibly kept
as they had been (or, in some cases, caused to undergo parallel or convergent alteration)
because of the advantageousness of their outwardly seen features, and in spite of those
genetic forces of mutation pressure, drift, and selection for other,
"correlated" characters, which would otherwise have succeeded in making them
different from what they are. And it may be recalled that, in many of
these instances, not merely a given character as such, but the exact type and grade of
development of it, shows this behavior, as in the case of the very precise bristle pattern
of the two Drosophila species cited.
The
attainment of very nearly the same end-results, as shown in some of their
characters, even by species of extremely different descent and genetic background, when
their mode of life and surrounding circumstances are similar, provides
even more striking evidence of the usefulness of these characters and of the precision
of selection in molding them, as many comparative anatomists, paleontologists and
other Darwinian evolutionists have for a long time recognized. Thus the very
similar fishlike, accurately streamlined form not only of widely different fish species
themselves, but even of the reptilian fossil Ichthyosaurs, as well as of modern whales,
bears eloquent testimony to this.
Another
oft quoted example is that of the carnivorous marsupial called the Tasmanian wolf, which,
living a wolf-like life, has became so wolf-like in its proportions and general appearance
as readily to be mistaken for one despite its much closer genetic relationship to the
opossum. Such illustrations could be greatly multiplied, especially if individual organs
or features, such as the independently evolved eyes of cephalopods, vertebrates and
scallops, were singled out. They prove that the significance of these characters for the
organism lies in the end-results themselves, -- i.e., in the advantages which the latter
as such confer, -- and that the genetic and developmental mechanisms whereby the results
arrived at are matters of comparative indifference, varying greatly with the "accidents" of mutation and of past history, that have worked
on a biological material which is essentially very plastic, at least from the standpoint
of morphology and proportions. This very plasticity would, however, soon result in its
becoming buffeted completely out of shape, in the course of less than one geological
period, were it not for the steadying hand of selection,
that persists in actively maintaining such a remarkable precision of genetic adaptation. |
|
Commentary:
Characters in allied species, living in similar habitats, may be preserved over millions
of years, yet if the two "species" can be crossed to produce hybrids, here the
characters (e.g. bristle pattern) may "show
considerable disturbance". Thus, while a "given character remained in ... its original form during the
evolution of both species, nevertheless the [hidden]
genetic and developmental basis beneath it underwent
changes" (as a product, and perhaps cause,
of speciation events). The "genetic basis had
become different, while maintaining a similar facade"
[conventional phenotype]. Muller here points to the
inheritance of genetic information at more than one level
.
|
INDIVIDUAL
VARIABILITY AS A MEASURE OF THE LATITUDE OF ADAPTIVENESS
Nevertheless it should be recognized that some species appear much
more variable that others, and that even different characters in the same species can show
considerable difference in the amount of individual variation that exists in them and in
the degree to which such variation seems to affect the welfare of the organism. Thus, to a
European man it probably makes very little difference in his expectation of life in Europe
whether his hair happens to be light or dark, sparse or thick, unlike what we have found
above to be the case in our Drosophilae. The question accordingly arises: why should there
be so much greater variation of characters, and apparently greater tolerance of such
variation, in some cases than in others?
In examining this
question it should, in the first place, be understood that, in a large, fairly panmictic
population which has long existed as such, the amount of variation found in any given
character is largely a function of two sets of factors: the frequencies of mutations of
different degrees affecting that character, and the average amounts of disadvantage
conferred by the mutations of different degrees.
In fact, if we
make the simplifying assumption, which is probably good enough as a first approximation,
that the average amount of disadvantage is proportionate to the amount of deviation, a
simple relation can be calculated. This turns out to be as follows (stating it directly in
words rather than in mathematical symbols): The amount of disadvantage caused by an
(arbitrarily chosen) unit grade of deviation is equal to the mean absolute deviation
arising by mutation in the germ cells in any one generation, divided by the total genetic
variance of the population in respect to the given character. We need not trouble the
reader with the rather simple mathematical derivation of our formula, but only point out
that it depends upon the facts that the total variance represents an
accumulation of the mutations that occurred throughout many generations, and that the
amount of accumulation of those mutations that manifest themselves will be inversely
proportional to their disadvantage, that is, to the rate at which they are weeded
out.
If, then, we were to
determine the mutation rate for the given character in such a population, we could find
approximately what grade of disadvantage a given grade of deviation from the norm
entailed. (Included in this reckoning of the disadvantage, however, would be that arising
from any correlated effects of the given mutant genes.) A second way of arriving at the
same result, of course, would be by direct observations of the relative viabilities and
fertilities of the different types, and the results of these two methods should check.
Unfortunately, quantitative studies of this kind appear not yet to have been carried out,
important though they would be in the attack on problems of evolution.
In the absence of
quantitative studies of these kinds, it is nevertheless already evident that most of the
mutations which are viable enough to play any considerable role in the formation of the
observed variance manifest themselves with a good deal higher frequency in the population
than that frequency with which they arise anew in any one generation. That is, they do
accumulate to a considerable degree, and the disadvantage of each individual one is, on
the average, fairly small. Nevertheless, this disadvantage does prevent their accumulation
beyond the observed amount, and it usually results, at any rate, in the average grade of
the character, the norm, being rather well defined and comparatively stable. Moreover, we
become aware of the falsity of the assumption so often made, by both biologists and
medical men, which holds that variants within the so-called "normal
range" (i.e. those falling within, say, the middle 80 or 90 per cent of the
area of the curve of variation) are in effect "normals,"
possessing no or negligible disadvantage.
Now although
differences in the mutation frequency per generation must undoubtedly exist between
different species and, in all likelihood even more, between different characters of the
same species, it is probable that differences in the degree of disadvantage of comparable
deviations from the norm are, on the whole, a much more important cause of the differences
in the observed variance of populations found in different cases. An especial reason for
this is that the mutation rate is itself responsive to some extent to selection, and that
it would tend to creep upwards, through accumulation of mutant genes that themselves
allowed mutations to occur more easily, to the point where it was held in check by reason
of the disadvantage thus entailed. Hence, an especially low variance is probably, in most
cases, an indication of an especially high disadvantageousness of deviates.
We are then justified
in asking, why should there be such considerable differences as the evidence indicates to
exist between different species, as well as different characters, in regard to the degree
of deviation that causes a given degree of disadvantage? In answer to this, many different
illustrations will no doubt present themselves to the biologist and the medical man, of
physiological, biochemical, morphological and ecological relations which require different
degrees of precision. However, there are certain kinds of situations which are in general
conducive to lower requirements of this kind, or to a poorer meeting of these
requirements on the part of the population.
One major
situation which would, temporarily at least, produce an increase in the allowed variation
of a character is that in which, for any reason, a character that was previously of value
has now through some change of conditions (instituted not too recently) had its
value diminished or possibly even reversed. A similar effect may be produced if there has
been a swift change in regard to what constitutes the optimum grade of the character.
All this is, of
course, equivalent to saying that selection has relaxed, or has set up a different
standard than the previously established norm. As a result, many variants that previously
would have been eliminated quickly are now allowed a longer lease, or may even be caused
to multiply actively, and during the disequilibration the variance will increase until a
new norm is approached more closely.
Applications of
this principle to the situation among civilized humans and domesticated animals, in
respect to many characters, are obvious. There are somewhat similar applications in such a
case as that of the sloth, a mammal which follows a mode of life in which a high level of
activity, temperature regulation, and the use of its wits, together with many features
connected with these, have largely lost their advantages. In all these situations, then,
we should expect to find an increase in the amount of manifested variation, as in fact we
do.
Despite the above
relationships, it seems probable that many variations which hitherto have usually been
regarded as valueless, especially such physical ones as distinguish human races that live
in different climates, will be found to have their functional aspects, or at least to have
had them, in the days when they became prevalent. And even characters which occur only as
individual variations within groups, sometimes have a usefulness that maintains them up to,
and keeps them down to, their given level.
Instances in
point are the cases of dimorphic mimetic butterflies, as pointed out by Ford, the cases of
inversions studied in some Drosophila species by Dobzhansky and by Dubinin, and, if I am
right, such a character as near-sightedness in man. The latter, according to the
interpretation arrived at independently by the present writer and by Riddell (both
unpublished), was probably of considerable aid, in primitive tribes, to the relatively few
specialists who fashioned arrowheads and did other fine work.
This kind of
process, then, makes for intraspecies variability, but in doing so actually increases the
precision of adaptation. Multiple genetic choice in regard to camouflage characters, to
hinder recognition of a species by predators, as we judge to be the case in Nabours'
grasshoppers, and, conversely, a high variability in pattern to promote the recognition of
individuals by others of the same species, as perhaps in killifish (Lebistes), and
in mammalian scents, voices and faces, would come within the same general category of
phenomena.
Another major factor
in variability is, as Wright has analyzed with such remarkable insight, the extent, degree
and duration of sub-division of a species into semi-isolated populations, and the
effective breeding-size of these local groups. The smaller the groups the more does
accident, unrelated to value, determine survival of one gene rather than another within
the group, and the less precise can be the adaptation of the mean, not to speak of the
individual. At the same time, in the large, these local experiments in variation should
operate to further the possibilities of evolution and of a more far-seeing adaptation of
the species as a whole, through giving a wider range for selection and recombination
between the differentiating groups.
Whatever the reason
may be, whether because of the accidents born of small numbers in local groups, or --
probably less often -- because a given rare character or combination of characters that is
useful has happened to become tied up with a variation that would otherwise be
detrimental, considerable upsets in the mean values of given characters do sometimes occur
in evolution. And these may cause a loss in the precision of adaptation previously
attained, even where the optimum or standard of selection for the given character may
still remain largely the same.
Evidence of the
occurrence of such events is to be found, for instance, in the presence of "repeat" or "duplicated"
sections in the chromosomes of the normal Drosophila, and even in its X-chromosome.
These must have altered the dosage of many genes at once, and, in the case of those in the
X, many of the delicate dosage-compensation mechanisms must have been
much disturbed simultaneously, yet these aberrations were not too disadvantageous to
survive and become established. Following such occurrences, however, the population must
have settled down to a prolonged process of repair and readjustment, that perhaps allowed
the aberration to be worked out to the net advantage of the species in the end. Thus,
there is not only the patient "whittling" process
in evolution. However, as judged by the mass of results previously reviewed, this process
must be so much the more usual procedure than the other that the precision of adaptation
finally attained thereby is on the whole far greater than most biologists have hitherto
realized. |
|
Commentary:
Here Muller tries to "have his cake and eat it". He is trying to show the
precision of genetic adaptation, but has to admit an enormous amount of variability in
certain situations, especially the role of chance variation and random drift as a force in
evolution.
|
SUCCESSES AND LIMITATIONS IN
ADAPTATIONS
To sum up in a nutshell the argument arising out of the
consideration of dosage compensation, the subject which has consumed the major portion of
our attention in this paper, we would say that this phenomenon is an illustration
of the fact that organisms, starting with similar needs but different genetic make-up --
that of the male and of the female, respectively -- nevertheless tend to develop the same
characters, even to the utmost degree of precision, in spite of the fact that, to attain
this end, they must evolve still more different genetic and developmental mechanisms.
For, since the female
has two doses of sex-linked genes and the male but one, special compensators must be
evoked, working to different degrees in the two sexes, that make the grand end-resultant
of these complex biochemical activities remarkably the same, to a degree far beyond the
limits of our unaided vision. This striking convergence in the effects attained in the two
sexes, despite the differences in the mechanisms of attaining them, a result which so
resembles that of purpose, can be rationally explained only on the ground of selective
advantage which, through elimination of the unfavorable attempts and multiplication of the
favorable, produces effects similar to those of conscious profiting by experience.
Essentially the
same kind of phenomenon is seen in the resemblances in character between two species, like
the case of bristle resemblance in Drosophila melanogaster and simulans, where
again the same end is attained by
means of demonstrably dissimilar genetic and developmental mechanisms, as is shown by the
upset in bristle formation in the hybrid between them.* [Footnote: See qualifications expressed in footnote on pages
214-215.] Again, it is as if a goal were aimed at,
regardless of the means of its attainment. And since this goal is in fact attained, it is
necessary to infer here too that it has had, and in all probability still has, a value for
the organism, and that to depart from it is disadvantageous.
It is therefore justifiable, in this sense, to regard the organism teleologically.
And since its adaptations are found to be so far-reaching and precise, we are entitled to look for a function or functions of some sort
in practically every feature found, which has attained widespread,
long-term existence in a species. Owing to the complexity and the reconditeness of many of
the features of any organism's physiology, biochemistry and ecology, however, the functions in question will often be far from obvious to our
limited observations and imaginations. However, we may usually assume
that, until we have found them, we have not yet properly done our job of understanding the
organism. In the meantime, this realization itself should often be of help to us in our
effort to understand it, for we may thereby be guided to see in which directions some of
the gaps in our knowledge still lie.
Thus, the very fact
that the organism can do without the tonsils or appendix should have served to stimulate
further search for the long unrealized functions of these organs. And the fact that in so
many common infections the temperature is raised should early have made us suspicious
that, despite the inconvenience thus entailed, an adaptive mechanism was involved here,
which could sometimes be taken advantage of, and guided, rather than nullified [say, by
taking aspirin]. At the same time it should also be remembered, wherever there is a
host-parasite or even a symbiotic relationship between two species, that a given
development in one, say the host, may, like a gall, represent an adaptation achieved for
the benefit of the other -- the parasite -- rather than of the host itself. And by a piling up and interlocking of the adaptations and counteradaptations in
such cases, the problems involved may become exceedingly intricate.
The above
illustrations will also serve to illustrate another point not previously mentioned here.
That is, that many "final" characters, like many
genes themselves and many intermediate processes of development and physiology, serve more than one function, and may even be, at the same time,
actually disadvantageous in other respects, but with the disadvantages outweighed and
perhaps in part compensated for by special means.
In other words, even when we seem to have found "the" function of a part or process, we
may be far from finished in our search since there may be still other more or less
important consequences of it for the life of the organism. In a sense, each part
represents a compromise between advantage and disadvantage, if only because of the fact
that simplicity is usually an advantage, for reasons of economy in construction,
maintenance and repair, and for the minimizing of hazards, while on the other hand a given
use may often far outweigh the drawbacks of the greater complication. This all goes to
indicate how tremendously interwoven and intricate the life
processes of any higher organism are, and how the weighing of the benefit
of a given feature by the criterion of selection may often lead to a result quite
different from what we might have guessed.
Of course all this evidence of adaptation does not connote that natural selection
has acted like an omniscient guide. It can seldom take steps that depart very far
from a purely immediate advantage, even though it is helped to a limited extent by the
somewhat more far-flung trials engaged in by small semi-isolated
local groups, as previously mentioned. But some developments, which would require
much longer, more elaborate trials before their advantage materialized, will never have a
chance to be carried out. As an illustration we may cite the absence of all wheel
mechanisms in organisms, occasioned by the fact that such an arrangement cannot be arrived
at by a series of small steps giving intermediate stages which themselves are viable and
useful.
Conversely, other
developments, which for some temporary reason confer an immediate advantage, will become
incorporated despite their lower, or even negative, long-term evolutionary value. So, for
example, "mistakes" are made like placing the optic
nerve fibres in the path of the light, at a time when the eye is still so imperfect that
this does not matter so long as the function of distributing nerves to the individual
retinal cells is achieved. Once incorporated, however, such an
arrangement cannot easily be revoked, since around it, as around a keystone in an arch, there have in the meantime been placed other structures, now
indispensable, which would collapse if it were changed, unless very special substitute
arrangements were found. This, then, is a type of developmental correlation which cannot
readily be overcome. |
|
[Comment:
Sometimes the "other structures" are just embellishments, as suggested
by Samuel Butler when writing of an alternative "water dipper" for
his paint brush, and by Gould & Lewontin
(1979) in "The spandrels of San Marco and the Panglossian paradigm: a critique of the
adaptionist program". Proc. R. Soc. B. 205, 581-598]
|
There is, however, no contradiction here involved to our earlier thesis that the
characters representing branched effects of the same gene can be modified separately from
one another. For in cases like the present one the characters in question are not caused
by different branches of one primary process, but by the direct dependence of one from the
other chainwise, without branching, -- i.e., in what is in effect a straight line. Thus,
in our example above, the only use we can now expect to find in the given peculiar
arrangement of nerve fibers is that it serves in us as a necessary ontogenetic basis for
the development of other features, in themselves useful.
Considerations
similar to the above apply to many other facts of recapitulation in general, e.g., to the
development of gill slits, mesonephros, etc., in human embryos. Despite this persistence
of such relics, however, the course of development itself (as well as of intermediate
physiology) can undergo adaptive changes, while leaving the final product relatively
unchanged, so that in the end entirely new developmental (or physiological) routes to the
attainment of given adult structures (or reactions) are sometimes arrived at. A striking
illustration of this is seen in the substitution of "complete
metamorphosis", with its inserted pupal stage, in higher insects, for the
earlier "direct" processes of development. But that
this substitution was itself gradual can also be seen, from the intermediate situation
still persisting in some forms.
The organism, then,
is fearfully and wonderfully adapted, to a marvelous degree of nicety, but it has been
constructed bit by bit, by accretion and patching, as one minute, functionally operating
step was superimposed upon another. And though no unchanged part of the original fabric
may any longer be present, much of the earlier, and now superseded, pattern still remains,
in some way overlaid.
Therefore it is
probable that much of the present structure and working is far more complicated than would
have been necessary to achieve the present results, if only they could have been planned ab
initio for the most efficient attainment of the ends which are subserved today. This,
however, would have required intelligence, involving long-range foresight and design of an
order far beyond any which we ourselves as yet possess. The adaptation of organisms, in
other words, is most meticulous and all-embracing, yet close scrutiny shows it most
wanting in those respects which would have involved distant prevision, rather than
short-range trial and error.
To be sure,
natural selection itself does attain longer range when choosing among much diversified and
widely separated groups, and operating over longer periods, but in so far as this happens
it finds itself more limited in its range of choice, since there are fewer groups to
choose between as the categories in question become more general ones. Moreover, they are
limited to presenting, for these higher processes of choice, or "courts of appeals", such patterns as were themselves developed
through the selection processes of shorter range.
Despite these
limitations on the operations of selection, the organism is understandable in its present
workings, and in its past origination, only if its ends
are taken into consideration, that is, if it is regarded not only "mechanistically" but also "teleologically",
in the sense previously set forth. It is the fact of its being so constituted as to work
toward ends, -- i.e., toward results favorable for its own multiplication and
perpetuation, -- that chiefly distinguishes an organism from an inanimate thing, even
though its operations are not guided by that actual consciousness of their ends by which
they so often seem to be.
However, this
also is the reason why, when what we call consciousness does at last enter the picture, it
too can be utilized by organisms in the further attainment of their ends. Unlike other
ends, however, these ends are at the same time beginnings, since the courses of their
lives are cyclic, with the difference from other cycles [being] that these multiply as
they proceed, and that they are subject to mutations which, if they serve to further these
ends, themselves become reproduced.
All the above will
perhaps help to make clear the essential correctness of the aphorism of the American
zoologist W. K. Brooks, to the effect that "The essence of life
is not protoplasm but purpose". We must, however, understand the term "purpose" here as involving only an analogy. If we do so, we
should have reopened to us many clues which the so-called mechanistic biologists
wastefully disregarded, when they started us on that otherwise promising course the strict
following of which has at last put us in the position of realizing that teleology, of a
sort, is a part of "mechanism" after all. |
|
Commentary:
It is "justifiable ... to regard the organism
teleologically". However, in order to do this
we have to have a clear idea what the "ends" are, and what possible ways organisms might have evolved to meet
these ends. The Kimura "neutralists", appearing after Muller, opposed his simplistic view. These web
pages argue that it is time to pay more attention to Muller's teleology.
|
SUMMARY
The attempt to interpret the structures and reactions of living
things, without reference to their adaptive significance, is a consequence of confused
thinking, stemming from the opposition to the Darwinian principle of natural selection. As
shown by illustrations from Drosophila, mutations provide abundant evidence of the
adaptiveness of existing "normal" features, as well
as material for determining how these features operate in furthering the life of the
individual or the species. Moreover, a study of the effects of changing the number
(dosage) of genes, and of the nature of the dominance of normal to mutant genes, leads to
the conclusion that not only is each normal feature of adaptive value, in a qualitative
sense, but that its exact grade of development, quantitatively
considered, represents the optimum value for it under the conditions that prevailed during
the period when it attained that value. For the activity of
each gene is usually so near its saturation level as to result in a high stability of its
effects, despite the presence of disturbing factors. It is this which ordinarily
results, secondarily, in its appearing to be dominant over its more weakly acting ("hypomorphic") mutant allele.
Beyond this, a study
of the expression of mutant genes which exist in two doses in the female and one in the
male (those in the X-chromosome), shows that special genetic mechanisms have been evolved for compensating for
the effects of these dosage differences, so as to make the amount of
effect in the two sexes much more nearly the same than it otherwise would be. The
mechanisms involve the presence of other, modifying genes in the X-chromosome,
that may be termed "compensators". These, in their
double dose in the female, reduce the activity of her two doses of the primary gene, so as
to make the effect like that of the one dose of primary gene and one of compensators in
the male.
It is
shown that actually these mechanisms must have been evolved for the function of equalizing
the effects of the normal genes rather than the mutants, even though we can seldom be
aware of this equalization in the case of normal genes, because of their above-mentioned
near-saturation level. In other words, the stabilization of expression attained through
the high level of activity of the normal gene does not by itself equalize the effects in
the two sexes sufficiently to satisfy the organism's needs, despite its giving effects in
one versus two doses, which are not to us perceptibly different. For the compensating
mechanisms which have been evolved serve to equalize them much more exactly. This shows
that differences in visible characters, which are of far finer grade than can be detected
by our own powers of discrimination, are nevertheless of adaptive significance in the
organism's struggle for existence, -- i.e., they are of selective value.
Experiments are cited
which show that these compensating reactions are no mere consequences of the sex
difference itself, that a given gene usually has a whole system of interacting compensator
genes, and that the systems for different genes are different from one another. Thus each
must have evolved through an accumulation of several or many steps, subliminal to our
perception. The second-order magnitude of this selection pressure is illustrated by
evidence of Lamy and the writer, indicating the incompleteness of the compensation for
genes in parts of the X-chromosome of D. pseudo-obscura which have
more recently been acquired by the X from another chromosome. The objection is considered that, instead of subliminal selection for the
characters under observation, the results may be due to the developmental correlation of
these characters with other, more important but unobserved characters, in regard to which
major differences were caused by the 1-dose: 2-dose relation of the sexes. It is
shown that this interpretation encounters serious difficulties, based on the lack of
completeness of developmental correlation. Thus, we are led back to the conclusion that
selection must, in fact, have dealt subliminally with the actually observed characters.
Further evidence of
the adaptive value of normal characters, and in fact of the value of the precise grade of
expression which they exhibit, is found in their frequent persistence virtually unchanged
over millions of years, and even in different species having similar needs. It is shown
that the frequency of mutation is high enough to have prevented such stability, had
selection not actively favored the given norm. The causes of deviations from high
stability are considered. A formula is given (p. 217) whereby the strength of such
selection can be calculated from the existing variability within a large, persistent and
interbreeding population, when the amount of variation arising by mutation per generation
has also been determined. Though data of the latter nature are as yet insufficient, it can
in the meantime be concluded that the prevalent assumption is incorrect which would regard
deviations within the so-called "normal range"
(e.g., within the central 8 or 9 deciles of a population) as being of negligible detriment
to individuals possessing them.
It is concluded that
long persistent morphological, physiological or biochemical features, characteristic of
large populations, should in general be assumed to have, or to have had, adaptive value,
under the conditions in which these groups evolved, and that the degree to which they are
normally developed is also of adaptive significance. Failure to find this significance, or
function, is to be taken as an indication of our ignorance, and of the need for a deeper
understanding of their workings. This justifies us in applying a "teleological" viewpoint here, which is in no wise mystical. In
medicine, it should, moreover, lead to caution in attempting the alteration of standard
bodily reactions, or the extirpation or alteration of structures normally present, until
the functions of these parts and processes, and of their normal degree of development,
have been well determined. At the same time, it must also be recognized that selection
itself has been subject to grave limitations. Some of these have been discussed above. The
organism is not perfect in any absolute sense, otherwise there would be no use in
medicine, or in any form of artificial "interference"
with nature. |
Commentary:
Muller believes he has shown
"that differences in
visible characters, which are of far finer grade than can be detected by our own powers of
discrimination, are nevertheless of adaptive significance in the organism's struggle for
existence, -- i.e., they are of selective value."
The following papers here argue that it is the
proteins encoded by the genes, acting
collectively as proteins per se,
not the characters they direct, which are the target of Natural Selection
(Forsdyke 1994). Furthermore, Muller's system is a special case, the
dosage compensation system. It
does not follow that, in general, small differences in visible characters, (such as in the pattern of shells observed by
Gulick in the Sandwich islands), have
arisen, or persist, by virtue of Natural Selection. To this extent the mutations which brought them about
appear "neutral", having been selected (isolated) by the extrinsic ("random
drift") or intrinsic influences favoured by Gulick, Delboeuf, Romanes,
Bateson and
Wright. While phenotypically
neutral, the mutations may be serving genomic forces (i.e. they may not be genotypically
neutral). |
REFERENCES
Brown, F. A. and Hall, B. V.:
1936. The directive influence of light upon Drosophila Melanogaster Meig and some
of its eye mutants. J.
Exp. Zool., 74:
205-220.
205-220.
Kalmus, H.: 1941. The resistance
to desiccation of Drosophila mutants affecting body colour. Proc, Roy. Soc. Lond. s.B, 130: 185-201. 185-201.
Kalmus, H.: 1943. The optomotor
responses of some eye mutants of Drosophila. J. Genet, 45: 206-213. 206-213.
Lieb, M.: 1946. The production of
I-IV translocations and a preliminary study of dosage compensation in Drosophila
melanogaster. M.A. thesis, Indiana Univ. 27 pp. (unpub.).
MacKenzie, K. and Muller, H. J.:
1940. Mutation effects of ultra-violent light in Drosophila. Proc. Roy, Soc. Lond. s-B, 129: Proc. Roy, Soc. Lond. s-B, 129:
491-517.
Muller, H. J.: 1932a. Some
genetic aspects of sex. Am.
Nat., 66: 118-138. 118-138.
Muller, H. J.: 1932b. Further
studies on the nature and causes of gene mutations. Proc. 6th Int. Congr. Genet, (Ithaca), 1: 213-255. 213-255.
Muller, H. J.: 1935. On the
incomplete dominance of the normal allelomorphs of white in Drosophila, J. Genet. 30: J. Genet. 30:
407-414.
Muller, H. J.: 1939.
Reversibility in evolution considered from the standpoint of genetics. Biol. Rev., 14: 261-280. 261-280.
Muller, H. J.: 1943. Evolutionary
trends. Address before Nat. Research Council Conf. on Common Problems of Genetics and
Paleontology, at Amer. Mus. Nat. Hist., New York, July 25, 1943. (unpub.).
Muller, H. J.: 1949.
Reintegration of the symposium on genetics, paleontology and evolution. Genetics, Paleontology, and
Evolution. (Princeton Univ.
Press), pp. 421-445. (Princeton Univ.
Press), pp. 421-445.
Muller, H. J., League, B. B., and
Offermann, C. A.: 1931. Effects of dosage changes of sex-linked genes, and the
compensatory effects of other gene differences between male and female. (Abstr.) Anat. Rec. 51 (Supplement): 110. 110.
Sturtevant, A. H.: 1929. The
genetics of Drosophila simulans. Carn. Inst. Wash. Publ. .
399: 1-62. [End
of Paper]

Supplementary references
not provided by Muller
Bridges CB (1922) The origin of variations in sexual
and sex-limited characters. American
Naturalist 56:51-63
Goldschmidt R (1928) The gene.
Quart. Rev. Biol. 3:307-324
Stern, C. (1929) Biol.
Zbl. 49,
241-290. [Stern is mentioned above, but not cited.]
|

